NÚCLEO Y CROMOSOMAS
Nucléolos: en ellos se sintetizan los ARN ribosomales, aquí se ubican los genes y codifican el ARN ribosomal para la formación de ribosomas.
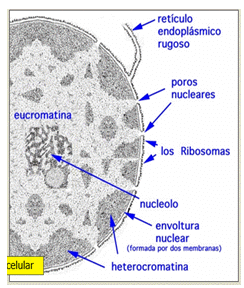
Cromatina: es la asociación de ADN y proteínas. Esta puede ser LAXA (o desarrollada, llamada Eucromatina, o COMPACTADA (llamada heterocromatina). si está compactada, el ADN no está disponible para que se transcribe (es decir, ser el molde para formar cualquier ADN). en cambio, si es laxa, se desencadena el proceso por el cual se genera ARN usando ADN como molde, proceso denominado TRANSCRIPCIÓN. la Eucromatina se ubica en el centro del núcleo, y es transcripcionalmente activa (disponible para transcribir). la heterocromatina es periférica y está anclada a la lámina nuclear; es una cromatina silente (es decir que no se genera ARN) y puede ser de dos tipos: CONSTITUTIVA (que es aquella que está compactada siempre, y se ubica en el centrómero de cada cromosoma), o FACULTATIVA ( es aquella que puede estar compactada e algunas células mientras en otras estará laxa, ya que todas las proteínas no son producidas en la misma célula).
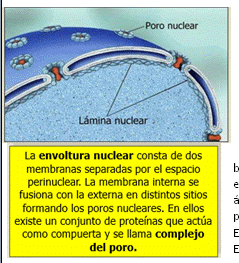
Lámina Nuclear: es una capa de proteínas dentro de la envoltura nuclear, la cual es una bicapa de fosfolípidos que forma poros y se ubica sobre la lámina nuclear. Los poros son una especie de caños con bordes de proteínas, por donde pasan moléculas muy grandes (como un ácido nucleico o proteínas), y salen aminoácidos o proteínas. Toda molécula que quiera entrar por el poro, debe estar relacionada con una proteína que lo reconozca, llamada IMPORTINA. En caso de querer salir por el poro, la proteína que reconozca a la molécula se llamará EXPORTINA.
COMPACTACIÓN DEL ADN
En las células eucariontes, el ADN se encuentra asociado a moléculas proteicas, las histonas, formando los nucleosomas, que actúan como unidad mínima de compactación de la cromatina. El nucleosoma está formado por un octámero de histonas rodeado por dos vueltas de ADN (una vuelta por cada par de 4 histonas).
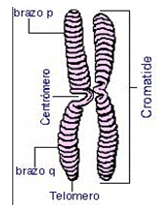
Los distintos niveles de empaquetamiento del ADN reciben nombres diferentes. El primer nivel, como ya se ha indicado, lo constituyen los nucleosomas. Están formados por las histonas H2A, H2B, H3 y H4. Alrededor de ese centro, se enrollan dos vueltas de ADN. Una quinta histona, la H1, sella la unidad y permite la conexión entre nucleosomas. El siguiente nivel lo constituyen los solenoides, que es que los nucleosomas vuelven a enrollarse, y se distribuyen alrededor de un eje imaginario. El nivel que sigue es el de las asas; éstos se superenrollan y forman, por último, los cromosomas, que consisten en dos moléculas de ADN unidas por una estructura llamada centrómero. Estos cromosomas están formados por una cromátida, lo cual es la misma estructura que posee un cromosoma simple con su centrómero (cromatina muy compactada) y sus brazos.
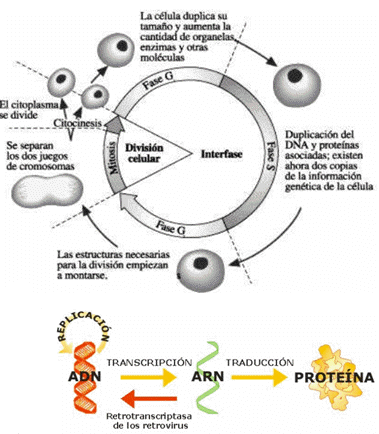
CICLO CELULAR
Las células, al dividirse, pasan por una secuencia regular de crecimiento y división celular: el ciclo celular (etapa de vida de una célula). Las primeras observaciones al microscopio óptico, determinaron dos periodos principales del ciclo celular: La división celular y la interfase. En la división celular se observa una gran actividad de reorganización y movimiento en la célula, donde el ADN se condensa haciéndose entonces visibles los cromosomas. La interfase, que comprende la mayor parte del ciclo celular. En esta parte, se duplica todo el contenido de la célula: tiene que duplicar el ADN, sintetizar más histonas (proteínas que se asocian al ADN), producir una cantidad de organelas adecuada para las dos células hijas y sintetizar las estructuras necesarias para realizar la división celular. Al iniciar la interfase, se tienen 2 cromosomas simple; luego de una serie de procesos, se duplica el ADN, del cual uno de ellos será el molde; a partir de allí, se unen dos cromosomas simples por su centrómero, formando un cromosoma doble, que seguirán el recorrido para llegar a la etapa de división celular.
CARIOTIPO
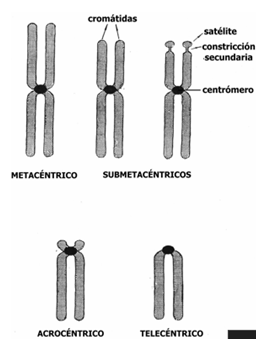
Es el número y morfología del cromosoma en una especie. La diferenciación está dada en el cromosoma nº 23, en el cual si la combinación es 2 cromosomas x, será de mujer, mientras que si se combina x e y, será de hombre. Consta de 23 pares de cromosomas. Los pares 1 a 3 son metacéntricos; el 4 y 5 submetacéntricos; el 6 a 12 y el X, submetacéntricos; el 13 a 15, acrocéntricos; el 16 a 18 submetacéntricos a acrocéntricos; el 19 y 20 submetacéntricos y el 21, 22 y el Y, acrocéntricos.
INTERFASE
Durante la interfase tienen lugar acontecimientos conocidos como “crecimiento”. El periodo de interfase se divide en las etapas G1, S y G2.
- Fase G1: es un período de intensa actividad metabólica. El tamaño de la célula se duplica, junto con sus organelas. Se sintetizan proteínas y lípidos.
- Fase S: la célula duplica todo su ADN y sintetizan solamente las histonas, que se unirán al ADN. Los cromosomas pasan de ser simples a ser doble (duplicación del ADN)
- Fase G2: es una fase en la cual la célula sintetiza proteínas relacionadas con la división celular.
SÍNTESIS DE PROTEÍNAS
Esta consiste en 3 procesos: TRANSCRIPCIÓN, PROCESAMIENTO (o maduración), y TRADUCCIÓN. El Dogma Central de la Biología decía que a partir del ADN, por transcripción, se forma la secuencia de nucleótidos de ARN. El ARN mensajero será el que lea y, por traducción, se unen los aminoácidos y se forman las cadenas peptídicas (replicación del ADN: se duplica). El dogma siguió vigente hasta que aparecieron los retrovirus, con la enzima retrotranscriptasa, capaz de fabricar ADN a partir de ARN.
Los genes son fragmentos de ADN que tienen la información para dar origen a un producto con una función celular específica. Están dispuestos uno a continuación del otro, separados por un espacio llamado Espacio Intergénico. La totalidad de los genes de un individuo o especie se denomina Genoma.
Las regiones del ADN son:
PROMOTOR: es el principio del gen; está ubicado en el 3’ del mismo.
SEÑAL TERMINAL DE TRANSCRIPCIÓN: es aquella parte que se ubica en el 5’ del gen.
TRANSCRIPCIÓN
El proceso de síntesis de ARN o transcripción, consiste en hacer una copia complementaria de un segmento de ADN (es un proceso anabólico y endergónico). El primer paso consiste en la asociación de la enzima ARN-polimerasa al promotor, para que forme una cadena complementaria y antiparalela a la que está usando como modelo (ADN). Es decir que la ARN polimerasa lee la hebra molde de ADN en sentido 3’-----5 ́y sintetiza la hebra de ARN complementaria en dirección 5 ́-----3 ́. Esto solo se realiza en esa zona, donde debe estar presente la eucromatina para ello.
La transcripción en eucariontes se lleva a cabo en el núcleo. Las células eucariontes poseen tres tipos de enzimas ARN polimerasas, que son ARN polimerasa I, II y III. Cada una de ellas se ocupa de la síntesis de diferentes moléculas de ARN. La ARN polimerasa reconoce al promotor del ADN por intermedio de proteínas llamadas factores de transcripción. Los ARN de transferencia (se pliega sobre sí mismo) y ribosomales (forma ribosomas) se transcriben de otros sectores del ADN. Los ARNm transcriptos sufren una serie de modificaciones antes de ser traducidos. Lo mismo sucede con los ARNr y ARNt.
MADURACIÓN O PROCESAMIENTO
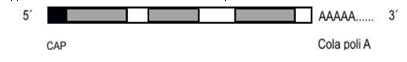
En células eucariontes, los diferentes ARNs son modificados antes de salir del núcleo y comenzar la síntesis de proteínas. El ARNm recién transcripto (ARNm transcripto primario) posee secuencias con información para la síntesis de proteínas, llamadas exones, y secuencias sin información, intercaladas, llamadas intrones. Este transcripto primario sufrirá varios procesos que lo modifican.
1) CAPPING: consiste en el agregado de un capuchón (cap) en el extremo 5 ́ del ARNm. El cap es un nucleótido con un metilo en la posición 7 de la guanina. Su función es impedir la degradación enzimática del ARNm inmaduro.
2) POLIADENILACIÓN: implica el agregado en el extremo 3 ́del ARNm de la cola poli A, lo cual son nucleótidos con base nitrogenada adenina.
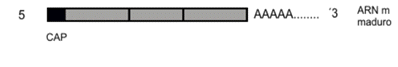
3) SPLICING: es el proceso en que los intrones son cortados, mientras se unen los exones, produciendo una molécula más corta que la inicial, llamada ARNm maduro. Para esta síntesis de la ARN polimerasa, se necesitan nucleótidos trifosfatados, a los que se le rompen dos enlaces para liberar energía que será utilizada en la formación de la unión fosfodiéster. El SPLICING ALTERNATIVO es la combinación de exones de distinta manera, cuyo objetivo es obtener diferentes mensajeros maduros, cuyo producto son cuatro proteínas distintas.
SÍNTESIS DE PROTEÍNAS O TRADUCCIÓN
La información que contiene el ADN se transcribe a los diferentes tipos de ARN. Los ARN sintetizados en el núcleo celular (en células eucariontes) son modificados antes de salir hacia el citoplasma, lugar donde se sintetizan las proteínas. La traducción es el proceso por el cual la información que contiene el ARN mensajero se utiliza para la síntesis de una proteína. Esta información determina el orden en que se unirán los aminoácidos, es decir, la estructura primaria de la proteína. El ARNt se forma como una sola cadena plegada en forma de hoja de trébol, con nucleótidos complementarios. Los ARNt poseen un anticodón, que es complementario a un codón (grupo de 3 nucleótidos, o triplete) del ARNm. La síntesis de proteínas incluye básicamente dos etapas: la aminoacilación o activación de los aminoácidos y la traducción propiamente dicha.
1-AMINOACILACIÓN O ACTIVACIÓN DE LOS AMINOÁCIDOS: El aminoácido se une en el 3p a su correspondiente ARNt por la acción de la enzima aminoacil- ARNt sintetasa. Dependiendo del anticodón, será el aminoácido que se unirá, siendo que el ARNt se una, al menos, a uno de los 20 aminoácidos usados en la síntesis proteica.
2- TRADUCCIÓN: este proceso de la se divide en tres etapas: la iniciación, la elongación y la finalización, que describiremos a continuación.
- Iniciación: En esta etapa se reunirán los componentes del complejo de iniciación. Este complejo empieza a formarse cuando la subunidad menor del ribosoma reconoce el cap y se une al ARNm en la zona próxima al codón de iniciación. Simultáneamente se acopla el ARNt que porta el aminoácido metionina, con gasto de GTP (este ARNt será el que reconozca luego al codón inicio del ARNm). La subunidad menor del ribosoma se deslizará sobre el ARNm hasta localizar el codón inicio AUG (que se complementará con el anticodón del ARNt iniciador cuya secuencia es UAC). Queda así establecido el marco de lectura (es leída en orden consecutivo, en grupos de tres nucleótidos). Finalmente se acopla la subunidad mayor del ribosoma, ensamblándose de esta forma el ribosoma completo y funcional. El ARNt iniciador queda ubicado en el sitio P del ribosoma.
- Elongación: Un ARNt cargado con el aminoácido correspondiente ingresa al sitio A del ribosoma, que está libre, y se acoplará por complementariedad de bases al segundo codón del ARNm. El aminoácido que estaba unido al ARNt iniciador rompe su unión con dicho ARNt con lo cual se libera energía. Esa energía es utilizada para que ese aminoácido se una al aminoácido que lleva el ARNt ubicado en el sitio A. Se forma la primera unión peptídica, reacción catalizada por la enzima peptidil transferasa ubicada en la subunidad mayor del ribosoma. Como consecuencia de todo esto, en el sitio P el ARNt queda sin aminoácido y en el sitio A está un ARNt que lleva los dos aminoácidos unidos. El ARNt “descargado” sale del sitio P e irá a activarse nuevamente, quedando así el sitio P vacío.
Ahora el ribosoma se desplaza tres nucleótidos (un codón) hacia el extremo 3 ́del ARNm. Como resultado, lo que antes estaba en el sitio A ahora pasa al sitio P y lo que estaba en el sitio P sale del ribosoma. Este proceso se denomina translocación y requiere el consumo de una molécula de ATP. Luego de la translocación, el sitio P vuelve a estar ocupado (ahora por un ARNt con la cadena de aminoácidos en crecimiento) y el sitio A a estar libre. Este proceso se repite sucesivamente codón tras codón.
- Terminación: Se produce cuando aparece uno de los codones de terminación (UAA, UAG, UGA) en el sitio A del ribosoma. En este momento un factor proteico de terminación se une al codón de terminación e impide que algún ARNt con otro aminoácido se ubique en el sitio A, se produce la hidrólisis de la cadena peptídica del ARNt en el sitio P, el ARNm ya leído completamente queda liberado y finalmente se separan las dos subunidades del ribosoma.
Una vez finalizada la síntesis de una proteína, el ARN mensajero queda libre y puede ser leído de nuevo. Una misma molécula de ARN mensajero puede ser utilizada por varios ribosomas simultáneamente y producir varias copias de la misma proteína. A estas estructuras donde un mismo ARNm está siendo traducido por varios ribosomas en forma simultánea se las llama polirribosomas (o polisomas).
Relación entre la secuencia de bases en el ARNm y las proteínas: CÓDIGO GENÉTICO
El código genético es como un diccionario que establece una equivalencia entre la secuencia de nucleótidos del ADN y la secuencia de aminoácidos de las proteínas. Cada aminoácido está codificado por tres bases nitrogenadas o tripletes. Sesenta y un tripletes codifican aminoácidos y los tres tripletes restantes indican la terminación del mensaje. Las características del código son:
-Es universal, pues está presente en casi todos los seres vivos conocidos. Solo existen algunas excepciones en ciertas bacterias.
-No es ambiguo, pues cada triplete especifica sólo un aminoácido.
-Hay varios tripletes para un mismo aminoácido, por ello se lo llama “degenerado”. Esta situación implica la existencia de tripletes sinónimos.
Diferencias entre la traducción en eucariontes y procariontes: En eucariontes, el ARNm es monocistrónico, es decir que codifican para una sola proteína, poseen cap, y la transcripción ocurre en el núcleo y la traducción en el citoplasma. En cambio, en procariontes, el ARNm es policistrónico, es decir que codifican para varias proteínas, no tiene cap, y su transcripción y traducción ocurren en el citoplasma sin ningún tipo de procesamiento del ARNm.
REGULACIÓN DE LA EXPRESIÓN GENÉTICA
1- Regulación de la expresión genética en procariontes:
La síntesis de enzimas está dirigida y regulada por algunos genes ubicados en un segmento del ADN llamado Operón. Un operón consiste en un promotor que gobierna la transcripción de varios genes.
- OPERON LAC: Uno de los ejemplos más conocidos es el operón lactosa. Para poder realizar la transcripción de los genes estructurales, el operador debe estar libre, es decir, sin represor. Lo que ocurre en ausencia de lactosa es que el represor está activo, es decir que se une a la lactosa e impide así que la ARN polimerasa se una al promotor. Como consecuencia no se produce la transcripción. En presencia de lactosa, esta se une al represor y produce en éste un cambio conformacional que lo inactiva, de manera que ya no puede unirse al operador. En estas condiciones la ARN polimerasa puede unirse al promotor transcribiéndose entonces los genes estructurales. Como la lactosa al unirse al represor lo inactiva y se pone así en marcha al operón, decimos que la lactosa actúa como un inductor.
- OPERON TRP: otro ejemplo es el operón triptofano. En presencia de triptofano, se produce la unión de la represora con la proteína, de forma que se convierta en activa y no se dé la transcripción. En cambio, lo que ocurre en ausencia de este es que la ARN polimerasa se une y se produce la transcripción. C El triptofano es reprimible, ya que inhibiéndolo, puede transcribir.
2- Regulación de la expresión genética en eucariontes:
Existen controles a nivel del ARNm (splicing y splicing alternativo); a nivel traducción; y a nivel post-traducción.
-Condensación del ADN en heterocromatina: las regiones de cromatina que están súper enrolladas no se transcriben.
-Secuencias y proteínas que controlan la transcripción: secuencias de nucleótidos en el ADN que pueden controlar (aumentando o disminuyendo) la tasa de transcripción de un determinado gen (cantidad en unidad de tiempo).
- Metilación: agregado de grupos químicos metilo (CH3) a la citosina. Cuantos más grupos metilo posea un gen, menor será su posibilidad de expresión.
- Factores de Transcripción: proteínas distintas de la ARN polimerasa necesarias para iniciar la transcripción.
MUTACIONES
Las mutaciones son cambios o variaciones en la secuencia de nucleótidos del ADN o bien en la cantidad de material genético que posee un individuo, que ocurren en la etapa S del ciclo celular. Aquellas mutaciones en el genoma (material genético-ADN) se producen porque el mensajero es quien arrastra el error, y se extiende al ADN. Estas pueden aparecer por errores en la duplicación o transcripción, como así también pueden ser producidas por agentes mutágenos. La enzima (ADN polimerasa) que copia al ADN (cromátidas hermanas) se puede equivocar, pero tiene la capacidad de corregirse. En cambio, la ARN polimerasa no tiene esta cualidad. Si esta última cae en un intrón o en el espacio intergénico, no pasa nada, no hay mutación. Distinto es el caso si cae en el exón, donde deberá analizarse para revelar si hay o no mutación.
Si los codones generados en la mutación son sinónimos (es decir que difieren en la última letra), la mutación se denomina SILENCIOSA, ya que no afecta en nada por codificar al mismo aminoácido que si fuese la original.
Si en vez de formar sinónimas, cambia la primer letra del codón, cambia el aminoácido, lo cual genera dos situaciones: por un lado puede resultar ser beneficioso, porque mejora la función, persistirá en la especie y se dará la evolución; por otro, puede ocurrir que sea desventajosa en el caso que genere una proteína que no cumpla sus respectivas funciones.
Otro caso de mutación es aquel en el cual puede producirse el cambio de un nucleótido por otro en la copia del ADN.
Si se elimina un nucleótido, la mutación se denomina DELECIÓN, si se agrega uno, se denomina ADICIÓN (o inserción) y en ambas se produce un cambio en el marco de lectura.
COMUNICACIÓN CELULAR
SEÑALES: pueden ser físicas (Presión, cambios de temperatura, etc.) o químicas (Hormonas (ligandos), factores de crecimiento, neurotransmisores, etc.- todas ellas son moléculas señal).
Diferentes formas de comunicación mediada por moléculas señal: existen diferentes formas de comunicación celular mediada por moléculas señal. En el caso de hormonas esteroideas y de la glándula tiroides (relacionado al sistema endocrino), estas moléculas no polares atraviesan la membrana y se unen a receptores intracelulares. Las secreciones pueden actuar cerca de la célula que la produjo (secreciones paracrinas) o a distancia transportadas por la circulación (secreciones endocrinas). Las hormonas pueden tener un origen proteico (REG) o lipídico (REL), estas son secretadas es decir que son células de exportación, por lo que se involucra al sistema vacuolar. Las hormonas se producen en glándulas, pero también una célula puede cumplir las funciones que desempeñan las glándulas (células glandulares) los neurotransmisores son fabricados en la neurona, dentro de vesículas; estos se unen al receptor, participan en la formación de polipéptidos. Otro tipo de comunicación es el contacto célula-célula, donde el ligando de una se une al receptor de la otra.
RECEPTORES DE MEMBRANA
Son glicoproteínas de la membrana que se unen a una molécula señalizadora o ligando, la que puede ser una hormona, un neurotransmisor, etc. Presentan tres regiones características: una para unión con el ligando, otra intra bicapa y otra citoplasmática. Se clasifican en:
- Acoplados a Canal o Ionotrópicos: receptores formados por diversas cadenas (subunidades) proteicas que forman un canal. La unión de la molécula señal o ligando, produce cambios conformacionales (apertura del canal) y flujo de iones.
- Acoplados a Proteína G (liga GTP): la unión del ligando altera su conformación y aumenta la afinidad por otra proteína de membrana: la Proteína G, con la que se unen. El GDP de la Proteína G se sustituye por GTP, provocando la separación del receptor y la unión con una enzima que cataliza la formación del Segundo Mensajero (intracelular), quien desencadenará una vía de reacciones intracelulares.
- Enzimáticos (actividad enzimática propia del receptor o de una enzima asociada): su dominio citoplasmático está acoplado a una enzima (fosfatasa) cuya acción cambia la conformación de una proteína.
Transducción de señales: la mayoría de las moléculas señalizadoras son secretadas por una célula y se unen a receptores de las membranas de otras. Esta unión provoca reacciones químicas en la célula receptora (célula blanco). Ocurre entonces una cadena de reacciones citoplasmáticas que transmite y amplifica la señal recibida en la superficie celular hacia estructuras intracelulares, originando una vía de respuesta. El proceso se conoce como transducción de señales. La mayor parte de las transducciones de señal implican una combinación de estos dos mecanismos: segundo mensajero y reclutamiento de proteínas Cada vía es una serie de proteínas distintas que operan en secuencia mediante cambios de conformación de la proteína siguiente.
La transducción finaliza cuando ocurre alguno de los siguientes fenómenos:
- eliminación del mensajero extracelular (o ligando)
- endocitosis del receptor
- degradación enzimática del receptor
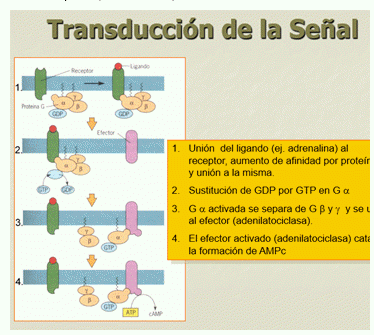
La secuencia de transducción de la señal es la siguiente:
- Unión del ligando (ej. adrenalina) al receptor, aumento de afinidad por proteína G y unión a la misma.
- Sustitución de GDP por GTP en G
- G activada se separa y se une al Efector (adenilatociclasa).
- El Efector activado (adenilatociclasa) cataliza la formación de AMPc
Rol del AMPcíclico: El AMPc (segundo mensajero) activa a la Proteína Kinasa A (PKA) que inicia una cadena de fosforilaciones. Existen numerosos procesos celulares que requieren del AMPc como activador de una vía metabólica. Por esto, muchas veces la cascada de reacciones requiere ser amplificada en el interior celular.
Amplificación de la Señal
1- La unión ligando-receptor activa varias proteínas G
2- Las AC producen varios AMPc a partir de ATP
3- Cada AMPc activa una PKA
4- las PKA activan moléculas de otra enzima
5- Las copias de la otra enzima producen muchas más moléculas de producto.
CICLO CELULAR
El ciclo celular se divide en dos periodos: La división celular y la interfase. En la división celular se observa una gran actividad de reorganización y movimiento en la célula, donde el ADN se condensa haciéndose entonces visibles los cromosomas. En la interfase, que comprende la mayor parte del ciclo celular, se observan diferentes procesos que determinan la secuencia ordenada de los preparativos que finalizan con la división celular. La interfase se divide en 3 fases consecutivas: G1, S y G2. Esta es la estructura básica del ciclo en células que se divide, pero no todas lo hacen. En aquellas dentro de este último grupo permanecen en un estado Go, el cual es una especie de “reposo” (ya que no es que no tienen actividad sino que no se dividen). El ciclo se detiene, para luego, volver a entrar a la etapa G1 y continuar el proceso.
La regulación y control del ciclo celular
Es conocido que, en cierto momento, las células normales cesan su crecimiento y se detienen en un momento tardío de la etapa G1 llamado punto R o punto de restricción. Una vez sobrepasado el punto R, las células completarán el resto del ciclo y finalmente se dividirán. El pasaje de la célula a través del punto R depende de la integración del conjunto de señales externas e internas que recibe. Otro mecanismo de control se lleva a cabo durante el proceso de duplicación del material genético, en la fase S, y asegura que la duplicación ocurra sólo una vez por ciclo. Luego, la célula entra en la fase G2 del ciclo. En G2, existe otro punto de control en el cual la célula "evalúa” si está preparada para entrar en mitosis. Este control garantiza que solamente entren en mitosis aquellas células que hayan completado la duplicación de su material genético.
En el control del ciclo celular intervienen complejos proteicos formados por dos tipos de moléculas:
1) Las ciclinas: son proteínas activadoras cuya concentración varía a lo largo del ciclo celular dado que alternan un período de síntesis creciente seguido por otro de rápida degradación. Constituyen la parte reguladora del complejo.
2) Las quinasas dependientes de ciclinas (Cdk): enzimas cuya concentración es constante a lo largo del ciclo y cuya activación depende de las ciclinas que así las regulan. Una vez activadas actúan fosforilando (agregando grupos fosfato) y activando como consecuencia a otras proteínas cruciales del ciclo celular. Las Cdk se activan cuando la concentración de la ciclina correspondiente alcanza su concentración máxima (esto ocurre cuando lo que se desarrolla en G1 sucede de forma correcta). Este es un requisito indispensable para que las dos proteínas puedan unirse y formar así un complejo proteico. Existen varios tipos de ciclinas que pueden agruparse en dos clases principales: las ciclinas de G1 y las ciclinas mitóticas. Las ciclinas de cada una de estas clases actúan en la fase correspondiente del ciclo celular.
Transición de G1 a S: A lo largo de la fase G1 va aumentando la concentración de la ciclina G1 hasta que la misma alcanza su umbral máximo. Como consecuencia la Cdk2 se activa y forma con la ciclina G1 un complejo denominado FPS (factor promotor de la fase S). La quinasa una vez activada fosforila a una serie de complejos proteicos relacionados con la duplicación del ADN lo que desencadena el inicio de la fase S. Dado que en un punto la concentración de la ciclina G1 comienza a declinar, se separa de la Cdk2 con lo cual el FPS desaparece. Esto permite controlar y asegurar que el ADN se duplique solo una vez por ciclo celular.
Transición de G2 a la división: La ciclina mitótica se acumula durante G2 hasta alcanzar su máxima concentración y se une a la quinasa Cdk1 formando el complejo proteico llamado FPM (factor promotor de la mitosis). La Cdk1 activa fosforila ciertas proteínas específicas, induciendo así los cambios estructurales que conducen a la mitosis. Todos estos eventos se revierten al concluir la división celular ya que las proteínas que los protagonizan se defosforilan al desactivarse la Cdk1, que ocurre cuando la concentración de la ciclina mitótica cae a un nivel inferior al umbral requerido.
¿Qué ocurre con el control del ciclo celular en células con daños en su ADN? Se ha observado en estos casos un incremento en la concentración de una proteína denominada p53. Dicha proteína actúa bloqueando el ciclo celular si el ADN está dañado. Este bloqueo otorga tiempo para intentar reparar el ADN. Si este daño es muy severo esta proteína puede desencadenar lo que se conoce como apoptosis (muerte celular programada). Si no hay alteraciones, los niveles de p53 son bajos, mientras que si ocurre el caso contrario, aumentan y conduce a la apoptosis.
REPLICACIÓN DEL ADN
Una de las etapas del ciclo celular es la fase S, donde la célula duplica su material genético. Esta etapa es crucial para la reproducción celular, ya que las células hijas deben recibir una copia exacta del ADN de la célula de cual provienen. Una vez determinado el ADN como el material hereditario y descifrada su estructura, los científicos comenzaron a investigar acerca de los mecanismos a través de los cuales el ADN se duplica.
¿Cómo se duplica el ADN?
Ya conocido el modelo estructural de la molécula de ADN, comenzaron a formularse hipótesis acerca de su duplicación, hecho fundamental que permite transmitir la información que contiene una célula a las dos células hijas resultantes luego de la división celular. Dentro de estas hipótesis, el experimento propuesto por Meselson y Stahl estos científicos demostró de manera concluyente que la replicación del ADN seguía el modelo de la hipótesis semiconservativa. En ella, la molécula bicatenaria de ADN se abre en forma de horquilla y se sintetizan dos hebras nuevas. Las hebras nuevas se forman, por complementariedad de bases, con la hebra “madre” u original que están copiando, lo cual asegura la preservación de la información genética. Al final del proceso se forman dos moléculas nuevas, formadas cada una de ellas por una hebra original (de la molécula vieja) y una hebra nueva. El nombre de “semiconservativa” proviene de que, en la molécula recientemente formada, se conserva una hebra de la molécula original. La duplicación del ADN presenta las siguientes características:
Semiconservativa: las moléculas nuevas conservan una de las hebras de la molécula original.
Bidireccional: a partir de un punto de origen, la molécula se duplicará en ambas direcciones.
Discontínua: la síntesis se produce a través de fragmentos en la hebra rezagada.
PROCESO DE REPLICACIÓN
Es similar tanto en organismos procariontes como en eucariontes. La replicación del ADN ocurre una sola vez en cada generación celular. Una vez que los mecanismos de regulación celulares disparan el inicio de la etapa S, comienzan a actuar sobre la molécula de ADN numerosas enzimas:
- Helicasa: rompe los enlaces puente de hidrógeno entre ambas cadenas complementarias del ADN, separando sectores de la molécula bicatenaria.
- Topoisomerasas: alivian la tensión de la molécula de ADN evitando su híperenrollamiento.
- ADN polimerasa: polimeriza los nucleótidos utilizando la molécula original de ADN como molde. Hay tres tipos diferentes de ADN polimerasas llamadas I, II y III. Entre éstas, la ADN polimerasa III es la responsable de la síntesis de ADN.
- ARN polimerasa o primasa: sintetiza pequeños fragmentos de ARN que se utilizan como cebadores.
- Ligasa: Cataliza la formación de enlaces fosfodiéster entre nucleótidos de distintos fragmentos (de Okasaki, ver más adelante).
La replicación comienza en sitios específicos llamados sitios de origen de la replicación (cadena molde), el cual es una secuencia de nucleótidos que será reconocido por la ADN polimerasa para hacer la copia de la cadena nueva. A partir de este sector actúan las enzimas helicasas separando ambas hebras de la molécula. De este modo se forma un área conocida como "burbuja de replicación". En los extremos, se genera una tensión capaz de romper el ADN. Aquí es donde intervendrá la topoisomerasa, que cortara la unión fosfodiéster de una de las hebras. Esta girara en sentido contrario, y se volverá a unir. Gracias a ese corte, alivia la tensión generada por la helicasa al romper las uniones puente de hidrógeno. Además, existe una proteína llamada SSB, que se coloca alrededor de la burbuja y estabiliza la forma monocatenaria de la hebra que se está replicando, para que no se pliegue y acople por complementariedad de bases con ella misma. A partir de esta acción, la ADN polimerasa es capaz de reconocer las bases libres y colocar el nucleótido con su correspondiente base complementaria (A-T y C-G). De esta forma se sintetiza una nueva hebra utilizando como molde cada una de las cadenas originales. La enzima ADN polimerasa III posee algunas restricciones para poder cumplir con su actividad catalítica. Una de ellas es que sólo es capaz de realizar la síntesis de ADN en el sentido 5´----3 ́, motivo por el cual opera en direcciones opuestas en cada hebra. Otra restricción de la enzima ADN polimerasa es que no puede iniciar la síntesis de las nuevas hebras si no cuenta con un extremo 3 ́ libre. Por esta razón es necesario que, antes del inicio de cada hebra nueva, actúe otra enzima, la ARN polimerasa o primasa, que sintetiza un corto fragmento de ARN que actúa como cebador. El ARN cebador o primer, corta la molécula de ARN y hace que empiece a actuar la ADN polimerasa (lo cual genera el antiparalelismo). Luego, la ADN polimerasa continúa la síntesis de ADN. Okasaki descubre que como la ADN polimerasa III solo puede ir en dirección 5’----3’, lo que hace es “saltar”, es decir que va construyendo las cadenas que comienzan en el 3’ en la dirección que puede pero cuando termina un fragmento se desplaza hacia el inicio de este y va formando otro, de forma que la síntesis global sea en sentido 3’---5’. Las hebras tardías o retardadas son aquellas que requieren del cebador, mientras que las hebras conductoras son las que van en el sentido 5’---3’. luego de todo esto, intervienen la ADN polimerasa I, que reemplaza a la primasa, con la ligasa, que une a la ADN pol I con lo que sigue de la hebra. La mitad de la burbuja se denomina horquilla. Para formar la cadena solo se necesita un fosfato, que se coloca como monofosfato, mientras que el sustrato es el trifosfato (o cuando están sueltos será di o trifosfato)
La diferencia en la replicación entre las células procariontes y eucariontes es que en ambas se siguen los mismos pasos, pero el origen de replicación es único en procariontes, y avanza bidireccionalmente hasta obtener dos moléculas de ADN circulares, cada una formadas por una cadena original y otra nueva (en eucariontes los orígenes son varios).
DIVISIÓN CELULAR
La Teoría Celular postula que todos los organismos están formados por una o más células y que todas las células provienen de células preexistentes. En este marco, se puede comprender la enorme importancia que tiene la reproducción celular para la continuidad de la vida en nuestro planeta. La reproducción de las células se lleva a cabo durante la etapa de División Celular del Ciclo de Vida de las células. Cuando una célula se divide transmite a sus células hijas dos requisitos esenciales para la vida:
- la Información Genética o información hereditaria para dirigir los procesos metabólicos.
- los materiales del citoplasma necesarios para vivir y utilizar dicha información.
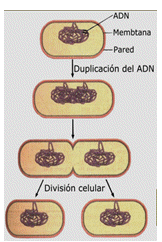
La división celular transmite un juego completo de material genético a cada célula hija
La información hereditaria de todas las células vivas se encuentra en el ADN. Como muchas moléculas biológicas grandes, una molécula de ADN es un polímero de subunidades llamadas nucleótidos. La unidad de la herencia, los genes, son segmentos de ADN de cientos o varios miles de nucleótidos de longitud, cuya secuencia constituye la información genética para producir tareas específicas. La secuencia de nucleótidos en un gen codifica la información para sintetizar el ARN y las moléculas proteicas necesarias para construir estructuras, y para llevar a cabo las reacciones metabólicas. Para que cualquier célula sobreviva, debe tener un juego completo de instrucciones genéticas. Por lo tanto, cuando una célula se divide, no puede simplemente dividir sus genes por la mitad y darle a cada célula hija la mitad de un conjunto. En lugar de eso, la célula primero duplica su ADN, y cada célula hija recibe, entonces, un "manual de ADN" completo que contiene todos los genes.
La división celular transmite a cada célula hija los elementos citoplásmicos esenciales.
Cada célula recién formada debe recibir las moléculas necesarias para leer sus instrucciones genéticas y conservarse viva el tiempo suficiente para adquirir nuevos materiales del medio y procesar nuevos componentes celulares. En general, cuando una célula se divide, su citoplasma se distribuye por igual entre las células hijas y este mecanismo proporciona a las células hijas todas las organelas, nutrientes, enzimas y otras moléculas que requieren. Las células recién formadas adquieren nutrientes del medio, producen sus propias estructuras y crecen. Después de un tiempo, dependiendo del organismo, del tipo de célula y de los nutrientes de que dispone, la célula se divide.
DIVISIÓN CELULAR EN PROCARIONTES
En condiciones favorables, los ciclos de vida de los organismos procariontes son mucho más breves que los de eucariontes. Luego de incorporar nutrientes y crecer, la bacteria comienza a duplicar la única molécula de ADN que posee. Como resultado de la replicación, quedan dos moléculas de ADN que se fijan a la membrana plasmática en puntos cercanos pero distintos, llamados mesosomas. La célula se alarga por crecimiento de la membrana plasmática hasta alcanzar el doble del volumen original, al mismo tiempo se invagina hacia el interior celular en la zona media entre ambas moléculas de ADN. Finalmente, la célula original termina por dividirse en dos células hijas; cada una recibe una molécula de ADN y la mitad del citoplasma. Con un juego completo de genes y con los materiales citoplasmáticos suficientes para metabolizar, cada célula hija crecerá y completará su propio ciclo. Esta forma de división se conoce como “fisión binaria”.
DIVISIÓN CELULAR EN EUCARIONTES
La división celular es similar en todas las células eucariontes. Consta de dos procesos: cariocinesis o división del núcleo, y citocinesis o división del citoplasma. Durante la cariocinesis la envoltura nuclear se desorganiza, y el material genético del núcleo, que ya se había duplicado en la etapa S, se separa en dos partes. Una vez separado, se forma una nueva envoltura en torno a cada parte. El resultado de la cariocinesis es, entonces, una célula con dos núcleos. En la mayoría de los casos, se divide luego el citoplasma, y el resultado final son dos células con un núcleo cada una.
Al proceso de cariocinesis se lo divide en etapas, las cuales son: profase–prometafase–metafase–anafase–telofase. Esta división en etapas es arbitraria, pero sumamente útil para detallar los procesos más importantes que tienen lugar durante la división.
MITOSIS EN LAS CÉLULAS ANIMALES
PROFASE
Al iniciarse la profase, la cromatina (ADN unido a proteínas) ya está duplicada debido al proceso en la etapa S. Por lo tanto, cada filamento de cromatina contiene dos moléculas de ADN idénticas entre sí. Cuando el núcleo se divide en dos, el material genético se reparte entre ambos núcleos hijos en partes iguales, y para ello, la cromatina, que durante la interfase se halla desespiralizada y en forma de filamentos, debe compactarse. Esto da como resultado las estructuras llamadas cromosomas, cada uno de ellos está formado por dos cromátidas hermanas unidas entre sí por una región llamada centrómero. Ambas cromátidas son idénticas, es decir, tienen la misma información genética; puesto que una es el duplicado de la otra.
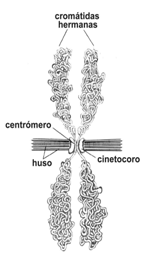
El centrómero está formado por regiones específicas de ADN de cada cromosoma. A ambos lados del centrómero, se forma una estructura proteica, plurilaminar, aplanada, llamada cinetocoro, cuyas zonas externas se unirán a los microtúbulos de las fibras del huso, fijando los cromosomas al mismo.
En un proceso posterior, que ocurrirá luego de la profase, las cromátidas hermanas se separarán, y cada una se desplazará hacia un extremo o polo de la célula. Para que eso se lleve a cabo, es necesario que en la profase ocurran dos acontecimientos:
- que se forme una estructura capaz de separar las cromátidas hermanas para transportarlas a los extremos de la célula
- que la envoltura nuclear se desorganice para que las cromátidas se puedan desplazar.
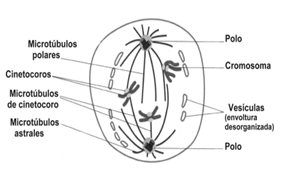
La estructura que separará a las cromátidas hermanas es el huso acromático, que comienza a formarse durante la profase y se completará en la prometafase. El huso acromático se forma en torno a los centriolos, estructuras cilíndricas formadas por microtúbulos de tubulina. Cuando la célula está en la etapa G1 de la Interfase, contiene un par de centríolos en el citoplasma, que se duplican en el período S, y dan origen a dos pares de centríolos. En la profase, cada par de centríolos comienza a migrar hacia cada extremo o polo de la célula y, mientras lo hacen, se organizan microtúbulos en disposición radiada en torno a ellos. A ese grupo de microtúbulos se lo denomina áster. A medida que se van separando, los microtúbulos van aumentando en longitud y en cantidad originando filamentos que en su conjunto, se denominan huso acromático.
Los microtúbulos están formados por unidades de tubulina, y aumentan de longitud porque continuamente se adicionan nuevas unidades de tubulina que se ensamblan. Este proceso de ensamble ocurre en una zona del citoplasma cercana a la envoltura nuclear, en torno a los centríolos, en un área poco visible al microscopio llamada centrosoma. El huso se va organizado por fuera del núcleo, entre los dos centrosomas.
Cuando comienza la división, el ADN que posee la información necesaria para la síntesis de ARNr y que forma el organizador nucleolar se espiraliza y enrolla, y, por lo tanto, el nucléolo se dispersa. En tanto, los cromosomas se hallan dispersos en el área que antes fue el interior del núcleo.
Prometafase: Se considera que comienza cuando la envoltura nuclear se desorganiza, formando una enorme cantidad de pequeñas vesículas, que permanecen cerca del huso durante toda la mitosis. El huso acromático ocupa, entonces, la región del núcleo. Los cromosomas se unen a las fibras del huso a través del cinetocoro. Los microtúbulos que participan de esta unión son los Microtúbulos del Cinetocoro. Los restantes microtúbulos que forman las fibras del huso se llaman Microtúbulos Polares, y, los microtúbulos que están por fuera del huso, alrededor de los polos, son los Microtúbulos Astrales.
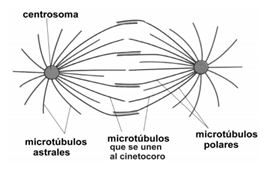
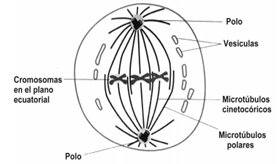
METAFASE
Al inicio de la metafase, la célula presenta un par de ásteres en cada polo y las fibras del huso, que se organizaron desde ellos, se encuentran distribuidas a lo largo de la célula.
Los cromosomas, unidos a los microtúbulos del cinetocoro, quedan entonces alineados en un plano, en la zona central de la célula, entre los polos, llamado plano ecuatorial.
Al terminar la metafase, todos los cromosomas se encuentran alineados en la placa ecuatorial y están conectados a los filamentos del huso.
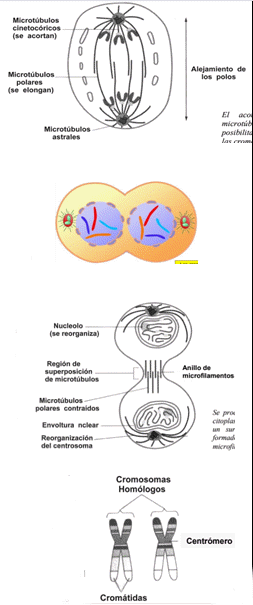
ANAFASE
En la anafase, las cromátidas hermanas, unidas por el cinetocoro, se separan, y cada una es arrastrada hacia los polos de la célula por las fibras del huso. Esto ocurre a partir de la separación de las placas del cinetocoro, que arrastran a cada cromátida hacia un polo. Las cromátidas migran debido al acortamiento de los microtúbulos cinetocóricos. Los microtúbulos polares, se elongan y los polos se van alejando. En esta etapa, el material genético se distribuye en partes iguales (serán el material genético de los futuros núcleos). Cada cromátida que migra es ahora un cromosoma formado por una sola molécula, ya que no está más unida a su duplicado. Dicho de otra manera, un cromosoma puede tener dos cromátidas (antes de la anafase) o estar formado por una sola (a partir de la anafase).
TELOFASE
Cuando los cromosomas llegan a los polos, el material genético ya se repartió y por lo tanto, la cromatina se desenrolla y adquiere la estructura que presenta durante la interfase. Al mismo tiempo, también se reorganizan la envoltura nuclear y el nucléolo. En esta etapa culmina la CARIOCINESIS, y el resultado final es una célula con dos núcleos con idéntica información genética.
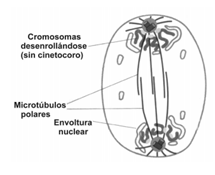
CITOCINESIS
Una vez obtenida la célula con dos núcleos, el citoplasma debe dividirse en dos porciones que darán lugar a dos nuevas células. Este proceso es la citocinesis. La división del citoplasma comienza a visualizarse con la formación de un surco alrededor de la célula, en el plano ecuatorial. El surco se distingue como si fuera un anillo en torno a la célula, que se profundiza cada vez más y estrangula al citoplasma, dividiéndolo en dos. El surco se forma debido a la contracción de microfilamentos de actina y miosina que se ubican por debajo de la membrana. Como resultado final, se obtienen dos células hijas que entran en el período G1 de la interfase. No en todas las células la cariocinesis es seguida de la citocinesis. Un ejemplo característico son algunos hongos, donde el núcleo se divide varias veces y se origina una célula con varios núcleos.
MEIOSIS: la División Celular como productora de Diversidad
Las células de todos los tejidos de un organismo, excepto las reproductoras, se llaman células somáticas. Si se examinan los cromosomas de una célula somática durante la metafase de la mitosis, se observará que cada cromosoma duplicado tiene un “gemelo” muy cercano que casi siempre es idéntico en longitud y posición del centrómero. En cada especie habrá un número diferente de cromosomas (en el humano, son 46), es decir, que el número de cromosomas es una característica propia de cada especie. En todos los casos, los cromosomas se observan en pares, y cada uno está formado por dos cromátidas hermanas unidas a la altura del centrómero. Los dos cromosomas de dicho par se llaman cromosomas homólogos, debido a que ambos portan genes que controlan las mismas características hereditarias. Por ejemplo, si el gen para el color de los ojos está localizado en un lugar en particular o locus (plural loci), en un cromosoma, entonces, el otro cromosoma del par (el homólogo) también tiene un gen para el color de los ojos en ese locus, a la misma altura.
Sin embargo, los dos cromosomas homólogos pueden tener versiones diferentes del gen para el color de los ojos, probablemente especificando colores diferentes. Los pares de cromosomas homólogos presentan dos tipos generales:
- un par lo constituyen los cromosomas sexuales, y determina, principalmente, el género de un individuo.
- el resto de los pares, los autosomas, determinan todas las características restantes.
En casi todas las especies de la naturaleza, las hembras tienen un par de cromosomas sexuales llamados cromosomas X. Por el contrario, los machos tienen un cromosoma X y otro Y. Estos dos cromosomas difieren en forma y tamaño. Sólo en pequeñas partes son homólogas; la mayoría de los genes que porta el cromosoma X no tienen a su contraparte en él Y. Sin embargo, las regiones homólogas son lo suficientemente grandes como para que los cromosomas X e Y se comporten como un par homólogo. Nosotros heredamos un cromosoma de cada par de nuestra madre y un cromosoma de cada par de nuestro padre, es decir, que cada uno de nuestros pares de homólogos consta de un cromosoma materno y otro paterno, formando dos juegos de cromosomas. Las células cuyo núcleo contiene dos juegos de cromosomas se llaman células diploides, y el número total de cromosomas o número diploide se simboliza como 2n. En los humanos, el número diploide es 2n = 46, pues poseemos 23 pares de homólogos en nuestras células somáticas.
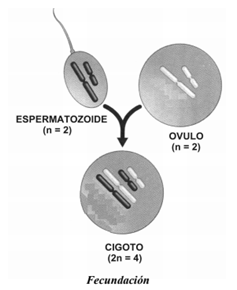
Las células reproductoras (óvulo y espermatozoide) se llaman gametas y cada una tiene un solo juego de cromosomas (en humanos: 22 autosomas más un solo cromosoma sexual X o Y), por esto se denominan células haploides. En los humanos, el número haploide es n=23. Las gametas se fusionan en el proceso de fecundación, dando como resultado una nueva célula llamada cigoto, que será diploide. El tener gametas haploides evita que el número cromosómico se duplique en cada generación. Las gametas se producen a partir de cierto tipo especial de división celular llamada MEIOSIS, la que se presenta solamente en los órganos reproductores (ovarios y testículos). Mientras que la mitosis produce células hijas con el mismo número de cromosomas que la célula original, la meiosis reduce el número cromosómico a la mitad.
Etapas de la meiosis: A diferencia de la mitosis, en la que ocurre una sola división que origina dos células hijas, en la meiosis se producen dos divisiones sucesivas. En la primera división meiótica se producen dos células, y cada célula hija volverá a dividirse, en la segunda meiosis, para dar dos gametas cada una. Una vez formadas, las gametas no se dividen y, si no participan en la fecundación, mueren. A su vez, cada división meiótica se divide en etapas que reciben el mismo nombre que en la mitosis, y muchos acontecimientos que ocurren durante ella son similares. Sin embargo, existen notables diferencias entre mitosis y meiosis, puesto que hay ciertos procesos que ocurren únicamente en la segunda. AI cabo de la meiosis II se obtienen, en total, cuatro células haploides.
MEIOSIS I
Profase I: AI igual que en la mitosis, el material genético de la célula ya fue duplicado en el período S. Al inicio, se visualizan los cromosomas al microscopio, pero no aparecen empaquetados como en la profase de la mitosis, sino como filamentos largos y finos, resultando difícil distinguir a las cromátidas hermanas dado que están muy juntas. A continuación, se produce el primer fenómeno distintivo de la meiosis: los cromosomas homólogos se acercan y comienzan a aparearse entre sí a lo largo. Ese apareamiento se denomina sinapsis, y es un apareamiento total, gen a gen de los homólogos. A Io largo de los cromosomas apareados se distingue una estructura formada por proteínas que es el complejo sinaptonémico, que contribuye a mantener a los cromosomas alineados. El apareamiento de homólogos es visible al microscopio óptico, y recibe el nombre de tétrada o bivalente.
Luego, se realiza un proceso que es la recombinación genética o crossing-over. El crossing-over consiste en el intercambio de segmentos de ADN entre los cromosomas homólogos apareados, y tiene por consecuencia que el cromosoma tenga ahora, en una de sus cromátidas, un segmento de ADN distinto al que tenía antes (tiene ahora la secuencia que estaba en su homólogo). Por Io tanto, las cromátidas hermanas ya no son idénticas entre sí. El complejo sinaptonémico desaparece y los cromosomas homólogos comienzan a separarse como si se repelieran entre sí, pero se mantienen unidos en Ios sitios donde hubo recombinación. Estos sitios se denominan quiasmas. Por último, los quiasmas se desplazan a lo largo de los cromosomas hasta sus extremos, en un proceso llamado terminalización. La profase I finaliza cuando la envoltura nuclear se desorganiza y el huso acromático comienza a formarse, tal como en la mitosis.
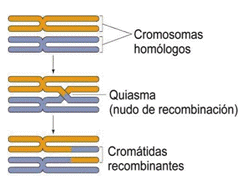
Metafase I: El proceso se realiza de manera similar al de la mitosis, exceptuando que los cromosomas ubicados en el plano ecuatorial, están apareados y unidos, aún, a través de las zonas de entrecruzamiento.
Anafase I: En la anafase I se separan los cromosomas homólogos, y cada uno migra hacia un polo de la célula, a diferencia de la mitosis, en la que migran las cromátidas hermanas. Los cromosomas homólogos no son iguales que al inicio de la meiosis, ya que durante el crossing-over intercambiaron fragmentos de material genético. Por otra parte, sus cromátidas hermanas no son idénticas puesto que una pudo haber hecho recombinación mientras que la otra no.
Telofase I: Los cromosomas homólogos llegan a los polos de la célula. En consecuencia, se tiene una célula con un número haploide de cromosomas en cada extremo, es decir, el número de cromosomas se redujo con respecto al de la célula original. Por esa razón, se dice que la Meiosis I es una proceso reduccional. Según la especie, puede o no formarse una envoltura nuclear en torno a cada grupo de cromosomas.
MEIOSIS II
La secuencia de procesos que ocurren durante la Meiosis II se asemeja a los de la mitosis, pero en este proceso, las cromátidas hermanas de los cromosomas no son idénticas debido al crossing-over, y que, además, las células hijas son haploides, no diploides.
- La profase II es breve, los cromosomas se vuelven a condensar, no hay apareamiento de homólogos porque solo está presente un cromosoma homólogo de cada par, si se formó una membrana nuclear se desorganiza y se establece el huso.
- En la metafase II los cromosomas, de dos cromátidas cada uno, se disponen en el plano ecuatorial de las 2 células, que son producto de meiosis I.
- En la anafase II las cromátidas hermanas no idénticas, unidas a las fibras del huso acromático por sus cinetocoros, se separan y migran a polos opuestos de la célula. Como en la mitosis, ahora cada cromátida se denomina cromosoma.
- En la telofase II en cada polo encontramos un miembro de cada par de cromosomas homólogos de una cromátida. Luego, se reensamblan las envolturas nucleares, los cromosomas se descondensan en forma gradual para formar hilos de cromatina y ocurre la citocinesis.
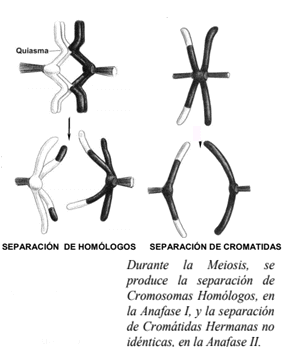
Las dos divisiones sucesivas de la meiosis producen cuatro núcleos haploides que presentan un solo cromosoma de cada tipo.
VARIABILIDAD GENÉTICA
En la meiosis, se producen células hijas genéticamente distintas a la célula madre, puesto que tienen la mitad del número de cromosomas, genéticamente diferentes entre sí, y, al mismo tiempo, son diferentes a las gametas producidas por otra célula madre. Las razones de esta variabilidad radican en el crossing-over, y en la migración al azar de los cromosomas homólogos y las cromátidas hermanas. Para ejemplificar, analizaremos una célula, de un organismo 2n=4, que se dividirá por meiosis, en la que los cromosomas oscuros son los heredados de la madre y los blancos, los paternos. En la anafase I, cuando migran los cromosomas homólogos, puede ocurrir que ambos cromosomas maternos se desplacen hacia un polo, o que un materno migre hacia un polo junto con un paterno y, el otro materno junto al otro paterno, migre hacia el otro polo.
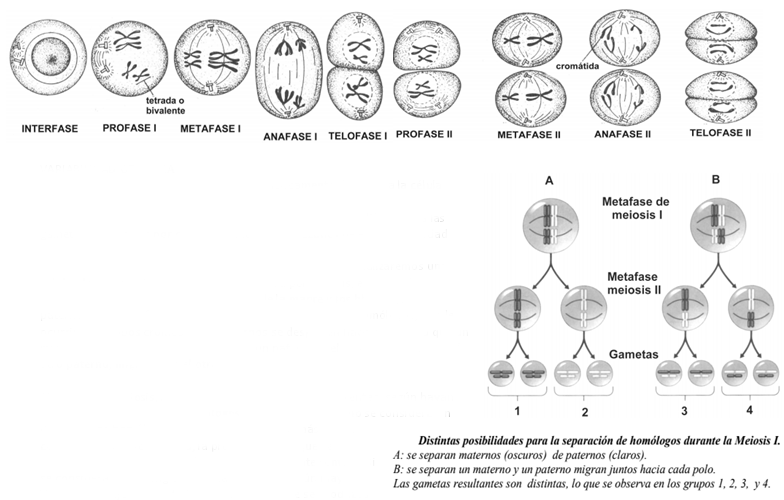
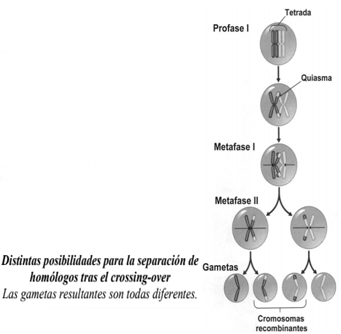
AI concluir la meiosis, las gametas resultantes serán diferentes, según hayan migrado los cromosomas homólogos. En este ejemplo, solo se consideraron dos pares de homólogos, pero si se consideran más pares, como en la especie humana, la probabilidad de que se produzcan dos gametas iguales resulta sumamente remota. Si se considera el crossing-over, la variabilidad es aún mayor. Volviendo al ejemplo anterior, supongamos que se produzca un crossing-over entre los cromosomas de un solo par de homólogos. AI igual que en el ejemplo anterior, supondremos que en la anafase I se dan ambas posibilidades: que ambos cromosomas maternos se desplacen hacia un polo, o que un materno y un paterno migren hacia un polo y el otro materno, junto al otro paterno, migren hacia el otro.
En este caso se producen cuatro gametas distintas en lugar de dos, como se habían producido cuando no hubo crossing-over en los casos A y B del ejemplo anterior. Como se puede apreciar, cuando se produce recombinación surgen nuevas variedades de gametas. Los ejemplos precedentes muestran que existen fuentes de variabilidad genética que originan gametas siempre diferentes entre sí. La primera fuente de variabilidad consiste en la migración, al azar, de los cromosomas homólogos durante la anafase I. Eso significa que cada cromosoma de cada par de homólogos migra hacia cualquiera de los dos polos, sin importar si es materno o paterno, o cómo migra el resto de los cromosomas. La segunda fuente de variabilidad es el crossing-over, proceso por el cual los cromosomas homólogos intercambian segmentos de ADN, y en consecuencia, las cromátidas resultan diferentes de las originales.
ERRORES EN LA SEGREGACIÓN DE CROMOSOMAS
Tanto en la anafase I como la anafase II, el material genético se reparte en partes iguales. Por ejemplo, en la especie humana, durante la anafase I se reparten 23 cromosomas (con dos cromátidas cada uno hacia cada polo de la célula); mientras que en la anafase II, se reparten las cromátidas hermanas, también 23, hacia cada polo. Puede ocurrir que, por algún error en el proceso de segregación, los cromosomas homólogos no se repartan en parte iguales, sino que migren 24 hacia un polo, y 22 hacia el otro. Este fenómeno se denomina NO DISYUNCIÓN. También puede suceder que la no disyunción ocurra en la anafase II, y que migre un cromosoma con dos cromátidas sin separar, en lugar de una sola cromátida. En el primer caso, se trata de una no disyunción primaria; el segundo, de una no disyunción secundaria. En ambos casos, el resultado será una gameta con un cromosoma de más. Si esa gameta es fecundada por una gameta normal, el nuevo individuo tendrá 47 cromosomas, es decir, uno de los pares de homólogos será un trío. En el ser humano, si el cromosoma adicional se encuentra en el par 21, el individuo presentará Síndrome de Down, enfermedad que produce retraso mental, rostro ancho, mayor separación entre los ojos, nariz aplanada, y otras características

MEIOSIS EN LA ESPECIE HUMANA
La meiosis se produce en las gónadas u órganos reproductores, es decir, los testículos y los ovarios. El proceso de producción de gametas se denomina gametogénesis. Más específicamente, espermatogénesis en el sexo masculino, y ovogénesis en el femenino. Aunque los procesos que ocurren en cada fase de la meiosis son iguales en ambos sexos, hay diferencias notables respecto de su duración y de la etapa de la vida en que se producen.
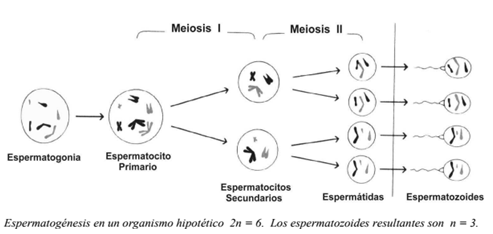
ESPERMATOGÉNESIS
En el hombre, la meiosis se produce en los testículos, a partir de ciertas células precursoras llamadas espermatogonias. Estas células son diploides y se dividen por mitosis, hasta que, en la pubertad, algunas se diferencian y se convierten en espermatocitos primarios. Estos serán las células que se dividirán por meiosis, y de la primera división meiótica, se producen dos espermatocitos secundarios. Posteriormente, cada uno de ellos se divide (segunda división meiótica) y originan en total cuatro espermátidas haploides. AI cabo de un proceso de maduración, las espermátidas se transforman en espermatozoides. El tiempo que transcurre entre la diferenciación de la espermatogonia hasta la formación de las gametas, es de unos dos meses.
OVOGÉNESIS
Los gonocitos se desarrollan a los veinte días de la gestación y, mediante mitosis, forman ovogonias (células precursoras). AI final del tercer mes de vida intrauterina, las ovogonias se diferencian en ovocitos primarios. Estos ovocitos primarios inician la meiosis para dar ovocitos secundarios, pero no la completan, sino que se detienen en la Profase I. En la pubertad, uno de esos ovocitos primarios reanuda la meiosis y se inicia el primer ciclo menstrual, ya que a partir de ese momento, un ovocito reanudará la meiosis cada 28 días. AI inicio de cada ciclo menstrual un solo ovocito primario comienza a aumentar de tamaño y reanuda la Meiosis I para dar el ovocito secundario. Paralelamente, las paredes internas del útero aumentan de grosor y se irrigan de vasos sanguíneos para alojar al futuro embrión en caso que el ovocito sea fecundado. A los catorce días desde el comienzo de la menstruación, el ovocito primario se libera del ovario maduro y concluye la Meiosis I. Esta primera división meiótica produce dos células hijas: una de ellas se llama cuerpo polar, la cual muere, mientras que la otra, llamada ovocito secundario, está en condiciones de ser fecundada. Si ese ovocito secundario es fecundado, se llevará a cabo la Meiosis II. AI igual que en la Meiosis I, una de las dos células hijas muere; mientras que la otra formará el óvulo.
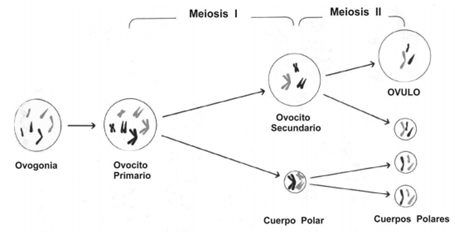
Una vez que el juego de cromosomas maternos se junta con el paterno, se forma el cigoto que se aloja en el útero. Si no hay fecundación, el ovocito muere a las 72 horas y los tejidos del útero se eliminan por la vagina durante la menstruación, y así culmina el ciclo de 28 días. A partir de ese momento comienza a madurar otro ovocito primario y se reanuda un nuevo ciclo.
Diferencias entre la gametogénesis femenina y masculina:
- En ambos sexos, la meiosis produce cuatro células hijas. Sin embargo, en el hombre las cuatro son viables, mientras que en la mujer solo una lo es, ya que los cuerpos polares no son aptos para ser fecundados.
- En el hombre, la meiosis se inicia en la pubertad, mientras que en la mujer, en el estadio embrionario.
- La meiosis en el hombre siempre tiene la misma duración. En la mujer, en cambio, puede durar entre doce y cincuenta y cinco años, según en qué momento se reanude la división meiótica de un ovocito.
- La mujer nace con un cierto número de ovocitos detenidos en meiosis, que en futuro darán origen a las células reproductoras. El hombre, en cambio, produce espermatocitos primarios a lo largo de toda su vida desde el inicio de la pubertad.
- La mujer libera una célula apta para ser fecundada cada veintiocho días aproximadamente. Por esto, solo podrá ser fecundada durante un determinado periodo. El hombre, produce millones de gametas continuamente, pero su fertilidad disminuye con la edad.
GENÉTICA
La genética, la ciencia de la herencia, es tema central entre las ciencias biológicas. Todos los seres vivos son producto de la interacción entre sus potencialidades biológicas y el medio ambiente en el cual se desarrollan. Por amplia influencia de la genética en todas las esferas de la vida, se ha convertido en materia obligada en una amplia gama de disciplinas científicas relacionadas con la salud humana y la producción agropecuaria. En una palabra, los seres vivos son siempre originados por otros seres vivos semejantes a ellos, pero esta semejanza no es absoluta ni incondicional. La semejanza entre padres e hijos no sólo depende de causas internas sino también de la relación de éstas y el medio ambiente. La genética moderna se originó cuando Gregor Mendel descubrió que las características hereditarias están determinadas por ciertas unidades que se transmitían de una generación a la siguiente de manera uniforme y predecible.
Cada una de dichas unidades, llamadas genes, son estructuras que deben cumplir, al menos, dos condiciones:
1) que se hereden de una generación a la siguiente de forma tal que cada descendiente tenga una copia física de dicho material,
2) que proporcione información, a los organismos que la portan, sobre la estructura, la función y otros atributos biológicos.
Como consecuencia de este doble aspecto del gen, hubo dos importantes enfoques históricos sobre los fenómenos genéticos: la identificación de la estructura química del material genético y el descubrimiento de la forma como se heredan los caracteres biológicos.
LA HERENCIA ANTES DE MENDEL
Los jardineros y los agricultores, acostumbrados a manejar y a cruzar variedades vegetales o razas animales, poseían un importante conocimiento práctico sobre el tema. Uno de ellos fue Thomas Makitta, maestro de escuela, quien enseñó la técnica de la hibridación en plantas con flores a algunos de sus alumnos interesados, entre ellos Mendel. Esos aficionados ya sabían lo que eran las líneas puras y los híbridos. Imaginen una planta que da flores rojas: si proviene de antecesores con flores rojas y además sus descendientes presentan tal color de flor, nos encontramos en presencia de una línea pura. Según la especie o la variedad vegetal de que se trate, se tendrán distintas líneas puras para los distintos colores de flores. En las plantas es posible la autofecundación, pues generalmente son hermafroditas. A partir del concepto "línea pura" es fácil llegar al de híbrido. Los jardineros y granjeros del siglo XIX conocían la importancia de la hibridación por cruzamientos sucesivos, que originaba razas y variedades intermedias de características reforzadas o potenciadas. Esas características podían ser benéficas o no, y el hecho de que aparezcan unas u otras todavía depende de la habilidad humana. Continuando con nuestro ejemplo, ¿qué pasaba si se cruzaban plantas de la línea pura "flores rojas" con plantas de la línea pura "flores blancas"? Los partidarios de la "mezcla" esperarían descendientes con flores rosadas. En algunas especies vegetales esto ocurre realmente así, pero en otras no. En éstas, los descendientes de un cruzamiento de ese tipo poseen, todos, flores rojas. Estas observaciones permitieron a Mendel enunciar una serie de Leyes que lograron explicar la mayoría de los fenómenos de la herencia. Se habla de tres leyes: la Ley de la Pureza de los Caracteres, la Ley de la Segregación al Azar y la Ley de la Segregación Independiente de los Caracteres.
LEY DE LA PUREZA DE LOS CARACTERES
A partir del cruzamiento de algunas plantas con flores rojas con otras de flores blancas o generación parental se puede obtener una generación de plantas con flores rojas o primera generación filial, o F1. Uno de estos individuos, por autofecundación, produce, a su vez, una generación de plantas con flores rojas y otras con flores blancas o segunda generación filial, o F2. En primer lugar, Mendel eligió como "sujeto" de experimentación la planta de arvejilla, y consideró siete caracteres hereditarios fáciles de observar, que presentaban siempre uno de dos rasgos alternativos. Con un criterio muy moderno para la época, Mendel analizó matemáticamente los resultados: calculó la proporción de individuos que presentaban cada rasgo estudiado y así obtuvo un patrón, una proporción constante que revelaba un orden interno. Vamos al grano (a la arveja): estudió de a uno por vez dichos caracteres y consideró a todos los descendientes, que en algunos casos eran varios miles. Mendel observó, por ejemplo, que si cruzaba plantas de semillas lisas con otras de semillas rugosas, toda la F1 (100%) presentaba semillas lisas, y que esta misma proporción se repetía cuando examinaba otros caracteres. La proporción se mantenía independientemente de que la característica a observar la portara el polen (gameta masculina) o el óvulo (gameta femenina) de la flor. Por todo esto, dedujo la primera ley, conocida como “Ley de la Uniformidad o Pureza de los caracteres”, que puede expresarse de la siguiente manera: “Al cruzar dos líneas puras, todos los descendientes del cruzamiento (F1), serán iguales en cuanto a la característica analizada”. Mendel determinó, entonces, que uno de esos caracteres o rasgos era dominante, es decir, se manifestaba en la descendencia, mientras que el otro era recesivo, es decir, quedaba "oculto" o no se manifestaba en la descendencia.
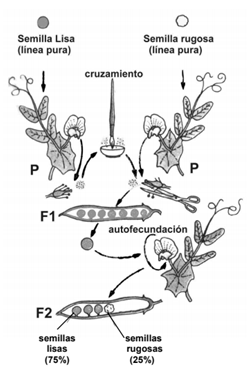
Representación simbólica y terminología moderna
Mendel propuso una hipótesis: Las características hereditarias estarían determinadas por factores dobles, de a pares, que se encontraban en todas las células y que se separaban al formarse las células sexuales. Por ejemplo, si se aplicara esta hipótesis al caso del color de las flores de la arveja, algunas de las plantas de líneas puras de la generación parental poseerían el factor "rojo-rojo", y otras, el factor "blanco-blanco", en una especie de "sitio" doble en el que se encontraría la información para el color de la flor. Las gametas de la arveja también tendrían un "sitio", sólo que no sería doble. La de las plantas con flores rojas incluiría el factor "rojo", y la de las otras, "blanco". Cuando, por ejemplo, una gameta masculina con el factor "rojo" se encuentra con una femenina con el "blanco", tiene lugar la fecundación y se forma un cigoto. Así, ésta tendrá, nuevamente, un "compartimiento" doble para el color de las flores, pero esta vez poseerá la información "rojo y blanco". Ese cigoto dará una nueva planta (F1), que tendrá las dos informaciones sobre el color, pero sus flores serán rojas porque ese rasgo domina sobre el blanco. Los individuos que poseen la información de los dos rasgos o características se denominan heterocigotas. Las líneas puras se conocen como homocigotas, es decir que poseen la misma información para una característica hereditaria. Las flores "rojo-rojo" son homocigotas dominantes, y las flores "blanco-blanco", homocigotas recesivas. La única ocasión en que puede llegar a manifestarse la información recesiva se da en el estado homocigota, cuando ningún otro factor la domina. Mendel supuso que cada característica está determinada por dos factores hereditarios, uno que proviene de la planta "paterna" y otro de la planta "materna”.
Propuso utilizar letras mayúsculas y minúsculas para simbolizar los factores dominantes y recesivos, y para facilitar los cálculos matemáticos en un papel. Así, designamos "R" al factor que determina el color rojo de la flor y "r" al que determina el color blanco. Entonces, de acuerdo con la información que poseemos para esa característica, podemos representar de la siguiente manera los distintos individuos: RR: homocigota dominante, flores rojas; Rr: heterocigota, flores rojas; rr: homocigota recesivo, flores blancas. Cada una de esas tres categorías recibe actualmente el nombre de genotipo. La expresión de los genotipos (flores rojas o flores blancas), fenotipo. A los factores hereditarios se los denominó luego genes, y a las alternativas posibles para ellos (R y r), alelos.
LA LEY DE LA SEGREGACIÓN AL AZAR
¿Se imaginan a Mendel en una mañana de mediados de 1860 plantando semillas de arvejas y siguiendo su germinación y desarrollo durante días a la espera de la floración? De la autofecundación de los individuos de la F1 Mendel obtuvo 929 semillas, que formaron la segunda generación filial, o F2. Cuando las flores mostraron finalmente su color, las contó y obtuvo 705 plantas con flores rojas y 224 con flores blancas. Entonces, Mendel calculó el cociente 705 / 224 y obtuvo la proporción 3,15 : 1.Podemos leerlo así: de cada 4,15 plantas, hay 3,15 con flores rojas y 1 con flores blancas. Esta experiencia se conoce hoy como cruzamiento monohíbrido, pues e trabaja con híbridos para un solo carácter (en este caso, el color de las flores). Mendel realizó el cruzamiento monohíbrido para las otras seis características, y en todos los casos, al calcular el cociente obtuvo siempre una proporción cercana a 3 : 1. Esto significa que, de cada cuatro individuos de la F2, tres presentaban el rasgo dominante, y uno, el recesivo.
A1 cruzar los individuos heterocigotas de la F1, cada uno de ellos aporta, en cuanto a la información que poseen para el color de las flores, dos tipos de gametas: uno "rojo" y el otro "blanco". Cada gameta tiene la misma probabilidad de encontrarse, en la fecundación, con una que presente el alelo para el color rojo o con una para el blanco.
Esto puede representarse en un tablero como el siguiente:

El interior del tablero muestra las combinaciones posibles de la información "rojo" y "blanco" -los distintos genotipos- en los individuos de la F2. Como se deduce de dicho tablero, de cada cuatro individuos, tres (un homocigota dominante RR y dos heterocigotas Rr) presentan el fenotipo dominante "color rojo", y uno (el homocigota recesivo rr), el fenotipo recesivo "color blanco". A menudo, este tablero es erróneamente interpretado: no indica que la F2 deba estar formada por cuatro individuos, sino que en la F2 se espera un 75% de descendientes con el fenotipo dominante de la característica estudiada y un 25% con el fenotipo recesivo. Pero esta probabilidad sólo se cumplirá en una F2 numerosa. Si la F2 está compuesta por sólo cuatro individuos, pueden resultar todos con flores rojas o todos con flores blancas, dos y dos, tres con flores blancas y uno con rojas, etc.
Estas características de la F2 llevaron a Mendel enunciar su segunda ley o Ley de la Segregación al Azar: “Los factores que determinan cada una de las características hereditarias, se encuentran de a pares en el individuo y sólo se segregan (separan) al formarse las gametas. Esta segregación es al azar. En la fecundación pueden unirse en nuevas combinaciones”.
LEY DE LA SEGREGACIÓN INDEPENDIENTE DE LOS CARACTERES
Medel consideró, luego, dos pares de caracteres: el color de las semillas (amarilla o verde) y su textura (lisa o rugosa). Para ello utilizó los siguientes símbolos: A: semilla amarilla, a: semilla verde; L: semilla lisa; l: semilla rugosa. Este cruzamiento se llama dihíbrido, pues se trabaja con dos caracteres simultáneamente: color y textura de la semilla. Mendel cruzó una planta de semillas amarillas y lisas con otra planta de semillas verdes y rugosas, ambas de línea pura (homocigotas). Al igual que lo que ocurre con la ley anterior, cada uno de los parentales produce un solo tipo de gametas, es decir, el individuo de genotipo AALL produce gametas AL, y el de genotipo aall, sólo gametas al. El genotipo de la F1 es AaLl, y el fenotipo, un 100% de semillas amarillas y lisas. Para obtener la F2, Mendel autofecundó a los individuos heterocigotas de la F1. Además del color, observó la textura de las 556 semillas que obtuvo de la F2. Las siguientes, son las cantidades de semillas en las cuatro combinaciones de fenotipos posibles:
Semillas amarillas y lisas: 315
Semillas verdes y lisas: 108
Semillas amarillas y rugosas: 101
Semillas verdes y rugosas: 32
Total: 556
En el siguiente tablero se representan las combinaciones posibles de gametas generadas por los individuos dihíbridos:
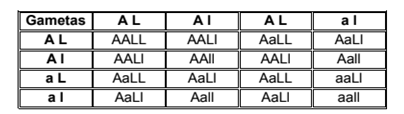
Si se divide cada uno de las cifras anteriores por la cifra menor (32) se obtendrá una proporción muy próxima a 9:3:3:1. Ante este resultado, Mendel se dio cuenta de que los factores hereditarios se separan cuando se forman las gametas y luego vuelven a unirse en la fecundación, lo cual produce nuevas combinaciones. Cada uno de estos caracteres se comporta como si fuera único, es decir, en forma independiente de los otros. Esto llevó a Mendel a formular la Ley de la Segregación Independientes de los Caracteres: “Cuando se forman las gametas, los dos alelos de un gen se separan independientemente de cómo lo hacen los otros dos alelos del otro par”.
DOMINANCIA INCOMPLETA Y CODOMINANCIA
Las experiencias muestran casos de dominancia completa: un alelo "domina" completamente al otro (recesivo), por lo que el individuo heterocigota presenta siempre el fenotipo dominante. Asimismo, se destacó que algunas plantas dan flores rosadas en la F1 como resultado del cruzamiento de dos líneas puras (rojas y blancas). En esto coincidían los partidarios "premendelianos" de una herencia mezcladora. Se trata de un caso típico de dominancia incompleta o intermedia, en la que el gen responsable del color de las flores posee dos alelos, uno para el color rojo y el otro para el color blanco. En el individuo heterocigota aparece el pigmento rojo en las flores, pero este alelo no domina totalmente al alelo recesivo blanco que llega a manifestarse parcialmente, y de ahí el color rosado de los híbridos. La codominancia es un fenómeno similar al de dominancia incompleta. La diferencia reside en que los alelos codominantes se manifiestan ambos por completo cuando están juntos, mientras que, en la dominancia incompleta uno de los alelos domina parcialmente al recesivo, permitiendo que éste se manifieste en parte.
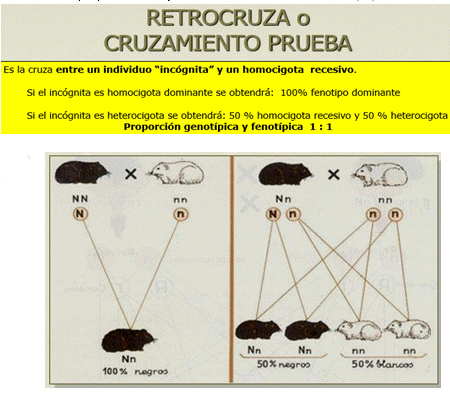
EL LIGAMIENTO DE LOS GENES
El genetista norteamericano Thomas Morgan (1866-1945) realizó estudios similares a los de Mendel pero utilizó a la mosca de la fruta. Una hembra normal de ojos rojos fue cruzada con un macho que presentaba ojos blancos. Toda la descendencia (F1) tuvo ojos rojos. Morgan interpretó que el alelo ojos blancos era recesivo y lo denominó “r” y al de ojos rojos, que era dominante, lo llamó “R” . Pero al continuar el cruzamiento y analizar a la F2, observó una proporción de fenotipos de 4 individuos con ojos rojos cada 1 de ojos blancos, en lugar de la tradicional proporción 3:1. Notablemente, todos los individuos de ojos blancos eran machos. Morgan cruzó al macho mutante original de ojos blancos con una hembra de ojos rojos de la F1. Obtuvo una F2 con iguales proporciones de machos y hembras con ojos blancos y ojos rojos: 1:1:1:1. Ante esto, propuso que el gen para el color de ojos se encontraba exclusivamente en un locus de un cromosoma al que llamó X. Asimismo, las hembras poseerían un par de estos cromosomas X, es decir, serían XX, en tanto que los machos serían XY. Así, Morgan hacía dos notables descubrimientos: por una parte, la existencia de cromosomas sexuales y, por otra parte, la ubicación de los genes en locus (lugar físico) de cada cromosoma, constituyendo grupos de ligamiento. Cuando los genes están muy juntos, se transmiten ligados (no se cumplen las leyes de Mendel), pero si se encuentran muy separados, lo hacen en forma independiente, cumpliéndose las leyes mendelianas. Un gen ligado al sexo es un gen cuyo locus está ubicado sólo en uno de los cromosomas sexuales, y está ausente en el otro.
HERENCIA LIGADA AL SEXO
Los genes se encuentran en los cromosomas y, para cada especie, hay un número fijo y específico de cromosomas. Todas las características determinadas por genes que se encuentran en el mismo par de cromosomas se denominan ligadas. En la especie humana uno de los primeros casos conocidos de ligamiento al sexo de un carácter específico fue el de desangramiento o hemofilia, enfermedad en que la sangre no coagula normalmente. Las experiencias de Mendel y otros científicos, habían demostrado que, en el macho, prácticamente todos los genes ligados al sexo, dominantes o recesivos, aparecen en el genotipo. Por eso los machos se consideran hemicigóticos para los alelos ligados al sexo. En las hembras los dos cromosomas X, proporcionan las relaciones de dominancia y recesividad, y se necesitan los dos alelos recesivos para que aparezca el fenotipo recesivo. Los machos heredan el cromosoma X de sus madres mientras que el cromosoma Y heredado del padre es prácticamente inactivo. En cambio, en las hembras reciben un cromosoma X de cada progenitor. (Daltonismo y calvicie)
EVOLUCIÓN
En la actualidad, los biólogos sostienen que todos los individuos pertenecientes a las distintas especies, descienden de formas ancestrales, por lo que todas las especies del planeta guardan un grado de parentesco. Desde el origen de la vida y a lo largo de millones de años, los seres vivos fueron cambiando, originando nuevas especies, así como extinciones. Este proceso continúa, aunque no es perceptible debido a la gran cantidad de tiempo que requiere. La “Evolución” es una rama de la biología que intenta explicar cómo se dieron y se dan estos cambios a lo largo de millones de años. A lo largo de la historia, hubo diferentes interpretaciones acerca del origen de las especies.
FIJISMO: En la antigüedad, se pensaba que las distintas especies fueron producto de una creación divina y eran inmutables. El fijismo es la postura según la cual las especies fueron creadas una única vez y nunca sufrieron modificaciones. Esta postura dice que los fósiles, animales extintos, no tienen que ver con el presente sino que simplemente desaparecieron.
CATASTROFISMO: Expone que la Tierra se creó hace 5000 años a.C., y que los cambios geológicos y la extinción de las especies están dadas por una serie de catástrofes.
UNIFORMISMO/UNIFORMITARISMO: Esta teoría dice que los cambios primero están causados por catástrofes y además pequeñas transformaciones continuas. En esta, se reconoce que en realidad el planeta Tierra era mucho más antiguo de lo que el catastrofismo decía.
TEORÍA DE LAMARCK: Lamarck formuló la primera teoría de la evolución biológica ya que propuso que los organismos cambian de acuerdo a sus necesidades, y esos cambios se van acumulando y transmitiendo a la descendencia. Los cambios en las especies están dados de forma “voluntaria” para adaptarse mejor al ambiente. Por ejemplo, según ésta teoría, un ave acuática desarrollará membranas entre las patas para poder desplazarse mejor en el agua.
TEORÍA DE DARWIN-WALLACE: Darwin partió del razonamiento de que los organismos producen más cantidad de descendientes que los que puede sobrevivir, ya que no hay suficientes recursos para todos. Esto da lugar a una competencia por los recursos entre los descendientes. Al mismo tiempo, todos los organismos son diferentes entre sí; pueden parecer iguales, pero no lo son, siempre presentan alguna mínima diferencia. Aquellos individuos que presentan características ventajosas con respecto a otros, tienen más probabilidad de sobrevivir y dejar descendientes, que a su vez, heredarán esas características ventajosas. De ese modo, los organismos van acumulando cambios graduales a lo largo del tiempo y así, a lo largo de millones años, se irían diferenciando en nuevas especies. Wallace llegó la misma conclusión: el mecanismo de selección natural es el motor de la evolución, permitiendo la supervivencia de los organismos más aptos para sobrevivir en cierto ambiente. Debido a los conocimientos de la época, Darwin no supo explicar el mecanismo de la herencia (cómo los caracteres se transmiten de padres a hijos) ni el origen de las variaciones entre los seres vivos. La selección natural es el proceso por el cual las condiciones del ambiente “seleccionan” determinadas características del individuo, permitiéndole sobrevivir o sobrevivir mejor que otro individuo que carece de ellas.
TEORÍA SINTÉTICA: Los conocimientos aportados por Mendel y los descubrimientos de la genética, permitieron explicar lo que Darwin no pudo: que las variaciones se deben a las mutaciones, al crossing-over y la segregación de homólogos al azar durante la meiosis, y que la información para las características de un individuo, se transmiten a través del ADN de generación en generación. La teoría de Darwin, complementada con los conocimientos de la genética, dio origen a la Teoría Sintética de la Evolución, enunciada a mediados del siglo XX. Con ésta, quedó descartada la teoría de Lamarck que había propuesto que los cambios beneficiosos adquiridos por los organismos durante su vida, se heredan. Esto significaría que un cambio fenotípico adquirido se codificaría luego en el ADN y luego se heredaría. Hoy se sabe que la información se encuentra en el ADN y a partir de ella se fabrican las proteínas que determinan el fenotipo, y que los cambios fenotípicos que persisten en la descendencia se deben a cambios en el ADN (mutaciones). La Teoría Sintética también dio lugar al estudio de la genética de las poblaciones. Esto fue así porque los individuos dejaron de ser vistos como el eje principal del proceso evolutivo, ya que éstos son elementos transitorios; puesto que lo que persiste es la población, no los individuos.
FUENTES DE CAMBIOS GENÉTICOS
Mutaciones: cambios.
Selección Natural: ambiente y adaptación.
Flujo génico: inmigración y emigración de las especies; una especie cambia de lugar de vida a otro, lo que genera cambios en la especie al cruzarse nativos del nuevo lugar con los inmigrantes.
Deriva génica: “cuello de botella” (debido a una catástrofe, se reduce la especie y se heredan las características que tienen los sobreviviente); “efecto fundador” (la migración genera nuevas especies en lugares diferentes al original, donde los primeros migrantes son los fundadores de esa nueva comunidad)
CONTROVERSIAS
GRADUALISMO: El gradualismo es la corriente de pensamiento que sostiene que los cambios profundos son resultado del producto acumulado de procesos lentos pero continuos, la cual rechaza la importancia de las catástrofes.
SALTACIONISMO: El saltacionismo es una corriente de pensamiento que propone que los cambios evolutivos son bruscos, ocurren de forma repentina y en gran magnitud, entre una generación y la siguiente. Ellos si aceptan la importancia de las catástrofes y también la teoría de Darwin, exceptuando el cambio gradual.