Semana 7
Teoría de Selección Sexual y Origen de los Sexos. Práctico.
En 1871 Darwin presentó el concepto de Selección Sexual el cual consiste en las características o rasgos aparecidos en distintas especies para aumentar las probabilidades de reproducción (ventajas reproductivas), por ejemplo, largas cornamentas, colores llamativos, etc.
El mecanismo de evolución por medio de la Selección Natural es aplicable a humanos. Rasgos heredables para los cuales existe variación, cambian sus frecuencias poblacionales si provocan variación en la eficacia biológica inclusiva de su portador, resultando en lo que conocemos como adaptaciones. Las adaptaciones surgirán a los saltos o continuas, rápidas o lentas, pero surgirán como producto de la Selección Natural.
El análisis de la Selección Sexual está caracterizado por una idea central: existe un conflicto de intereses entre machos y hembras durante la reproducción, donde cada sexo trata de maximizar su éxito reproductivo (E.R.).
SELECCIÓN SEXUAL
Origen y Consecuencias de la Selección Sexual
A) Origen de dos sexos: macho y hembra.
La reproducción sexual implica la formación de gametas a través del proceso meiótico y launión de dos gametas para dar origen a la cigota. Según existan o no diferencias entre dichas gametas se habla de sexo anisogamético o isogamético.
La anisogamia se diferencian dos sexos, macho y hembra. La gameta femenina (óvulo) es generalmente grande, con reservas de nutrientes y poca movilidad; la masculina ( espermatozoide) es por el contrario pequeña, sin reservas y muy móvil. Como tanto macho como hembra parten de cantidades semejantes de material gonadal (tejido para producción de gametas), aquel sexo que produce gametas grandes podrá generar pocas (hembra), mientras que serán muchas las que produzca el sexo con gametas pequeñas (macho).
El origen de la anisogamia se explica con la hipótesis de Parker, Bateman y Smith.
1) La reproducción es sexual, con producción de gametas por meiosis seguida de la fusión de gametas para formar el cigoto.
2) La probabilidad de supervivencia de un cigoto se incrementa en función del tamaño del mismo.
3) El tamaño total de las gametas producidas está limitado a la masa de tejido germinal (gonadal). A partir de dicha masa de tejido se pueden producir muchas gametas pequeñas o pocas gametas grandes.
4) Hay un tamaño mínimo de gametas por debajo del cual las gametas no son viables.
La hipótesis de Hurst y Hamilton está basada en la idea del "Conflicto Intragenómico" resultado de la acción de elementos genéticos egoístas. Durante la fusión de las gametas, el ADN nuclear de cada una de las células se junta y pasa a ser el nuevo material genético de la célula hija, pudiendo existir conflicto entre la expresión de la información contenida en la porción paterna y materna del nuevo núcleo. Pero donde se hace más marcado el conflicto es entre organelas como las mitocondrias (animales) y cloroplastos (vegetales), portantes estas de la herencia materna (ambos tipos de organelas poseen información genética pero que se transmite a través del citoplasma materno). La principal razón sería que los genes citoplasmáticos provenientes de organelas maternas y paternas no se juntan, sino que permanecen en organelas separadas y no hay razón por la cual deban compartir el citoplasma unas con otras. Debido a esto podrían surgir organelas mutantes capaces de eliminar a la "invasora", existiendo de hecho buena evidencia de que las mitocondrias pueden usar enzimas de restricción para degradarse unas a otras.
Los organismos que poseen sexo pueden ser divididos en dos grupos: aquellos que fusionan células durante el apareamiento y aquellos que se reproducen sin fusión y donde solo existe un pasaje de núcleo entre una célula y otra.
B) Consecuencias de la anisogamia y proporción de sexos:
1. Éxitos reproductivos diferenciales entre sexos: consecuencia de la anisogamia es que la proporción de gametas masculinas respecto de las femeninas se incrementa ya que el macho produce infinidad de gametas pequeñas y la hembra unas pocas grandes.
2. Proporción de sexos en relación a la selección sexual: Si cada macho está capacitado para fecundar gran cantidad de hembras, la competencia para acceder a parejas dependerá en gran parte del número de hembras que existan por cada macho.
Bateman comprobó estas teorías con una experiencia con moscas de fruta (Drosophila sp.) Bateman colocó igual número de machos y hembras (1:1) en un ambiente óptimo para la reproducción. A través de marcadores genéticos pudo individualizar los padres de cada nuevo juvenil y calcular así el éxito reproductivo de cada ejemplar. Los resultados demostraron que los machos tuvieron una mayor variación en el éxito reproductivo que las hembras; las hembras lograron en su gran mayoría aparearse y poseyeron éxitos reproductivos semejantes, esto esencialmente debido a que todas tuvieron acceso a un medio con abundancia de recursos para la producción de cigotas viables; La frecuencia de cópula (más allá de la primera) no tuvo efectos sobre el éxito reproductivo de las hembras. Los machos, por el contrario, cuanto más cópulas experimentaron mayor fue su éxito reproductivo. Se denominó el Efecto Bateman y el cual explica en líneas generales las estrategias reproductivas de uno y otro sexo.
La mayoría de las poblaciones de distintas especies en estado natural presentan una proporción de sexos de 1:1. Fisher explicó dicha proporción en término de la selección individual. Así, la proporción de sexos de una generación afecta el éxito reproductivo de esos individuos en la generación siguiente.
Cuando un sexo se convierte en un recurso limitante para el otro, el resultado consiste en un incremento en la competencia entre los miembros del sexo accesible para aparearse con ejemplares del sexo limitante.
C) Consecuencias de la inversión parental relativa:
El rol de los sexos: Competencia entre machos y hembras selectivas.
En las especies donde las crías reciben cuidado por parte de sus progenitores debe tenerse en cuenta la inversión parental. Esto sería lo que se hace para incrementar la supervivencia de una cría y que reduce la habilidad para producir una adicional. El progenitor que realice la mayor inversión parental estará reduciendo su capacidad para incrementar el éxito reproductivo futuro y por lo tanto existirá un conflicto entre padres en el momento del cuidado de la cría.Trivers relacionó la inversión parental con la competencia sexual y propuso que, el sexo que invierta menos en la cría competirá por parejas mientras que el sexo que más invierta será selectivo en aceptar parejas. Los machos compiten entre sí para conseguir la mayor cantidad de oportunidades para aparearse, mientras las hembras adquieren un papel no competitivo, seleccionando entre los machos cortejantes.
La anisogamia es el primer ejemplo de asimetría en la inversión parental de los sexos ya que los nutrientes que aporta uno y otro a través de sus gametas son diferentes y sesgados hacia una mayor inversión por parte de la hembra.
D) Selección sexual y evolución de los caracteres sexuales secundarios
La selección sexual es el proceso que modela los rasgos que dan a ciertos individuos ventajas sobre otros de su mismo sexo para competir y obtener apareamientos exitosos. Si la aparición de un "arma" (carácter utilizado para competir) le confiere ventajas a un individuo, se seleccionará la misma ventaja en sus competidores. Aquellos descendientes que hereden el rasgo serán, más exitosos, no significa que el rasgo se modificará en los sujetos existentes.
La intensidad de la selección sexual depende del grado de competencia por parejas. Esto está sujeto a dos factores: la diferencia en la inversión parental que realiza cada sexo y la proporción de machos y hembras reproductivamente aptos (llamada proporción operacional de sexos). Cuanto menor es la coparticipación de los sexos en la crianza, mayor es la selección sexual, debido a que uno de los sexos (el que menos coopera) dispondrá de tiempo y energía suficiente como para buscar nuevas parejas por las cuales deberá competir. Cuanto mayor sea la sincronización de ambos sexos en ser reproductivamente receptivos, menor será la competencia y por ende la selección sexual. La monopolización de parejas resulta energéticamente muy costosa debido a que en un corto tiempo se debe defender y copular a todas las parejas potenciales.
Los rasgos surgidos por la acción de la selección sexual son llamados caracteres sexuales secundarios y cumplen dos funciones principales, según actúen intrasexualmente o intersexualmente. Por un lado, para la lucha entre individuos de un mismo sexo y hacen a ciertos individuos más aptos para la competencia ( selección intrasexual). Por otro lado, actúan como estímulos discriminantes durante la elección que realiza el otro sexo y hacen que ciertos individuos sean más atractivos ( selección intersexual). Ejemplos de caracteres sexuales secundarios son: tamaño, agresividad, ornamentas, coloración, etc. y son reconocibles por cuatro propiedades básicas: (1) Están restringidos a un sexo (generalmente machos). (2) No se desarrollan hasta alcanzar la madurez sexual. (3) Generalmente, aunque no siempre, aparecen durante la época reproductiva. (4) Usualmente no incrementan la supervivencia, sino que la reducen.
Selección intrasexual.
Dos son las formas de competencia descriptas para machos, la competencia por parejas y la competencia espermática.
Competencia por parejas.
Se presenta en tres formas posibles:
A) Interacción agresiva que limita el acceso de otros machos a un recurso que las hembras necesitan o bien a las hembras propiamente dichas.
1. Los machos defienden recursos esenciales para las hembras. Cuando los recursos están distribuidos en forma impredecible o espacialmente agrupados, ciertos machos pueden defender esas áreas de mejor calidad. Si son escasas y críticas para la reproducción de las hembras, estás se concentrarán en dichos territorios y los machos dueños de los mismos se verán favorecidos.
2. Los machos defienden a las hembras como recurso limitante. Las causas del gregarismo en la hembra pueden ser varias, como por ejemplo el aumento de la eficiencia en la alimentación por estar en grupo, transmisión cultural de hábitos aprendidos, etc.
B) Competencia para dispersarse y encontrar hembras receptivas.
C) Competencia durante el cortejo para ser elegido por las hembras. Durante la época reproductiva los machos son sumamente activos y conspicuos.
Consecuencias de la selección intrasexual:
La consecuencia más inmediata es el acentuado dimorfismo entre sexos, debido al surgimiento de los caracteres sexuales secundarios. La selección intrasexual posee efectos sobre la supervivencia de los machos. Muchos de los caracteres sexuales secundarios modelados por dicha forma de selección, como son el incremento de tamaño, la producción de sonidos, la riqueza de colores, las armas para la lucha (cornamentas), etc., exponen a los machos a grandes riesgos de depredación y estados de estrés energético .
Otra consecuencia es un retardo en llegar a la edad en que son los machos reproductivamente aptos. Esto se debe principalmente al tiempo de maduración de las estructuras sexuales secundarias sin las cuales el individuo no está capacitado para competir por parejas.
Competencia espermática y Riesgo de paternidad.
El macho no sólo debe conseguir pareja; asegurarse que sus cópulas se conviertan en óvulos fertilizados es una de las tareas complementarias más importantes que debe realizar. Para ello, además de conseguir pareja y copular, debe evitar que la hembra copule con otros machos, y en caso de que esto ocurra, debe competir con los machos intrusos para que su esperma sea la que llegue al óvulo. Los machos más competitivos serán aquellos que produzcan esperma más móvil (menor tamaño) y de menor mortalidad.
Selección Intersexual
Darwin propuso que las hembras tenían ciertas preferencias en la elección de los machos basadas en caracteres estéticos de los mismos. Las críticas hacia la idea de Darwin se concentraron en que el fenómeno que proponía implicaba un desarrollado sentido de la estética por parte de las hembras. Otra fue que Darwin sólo estaba proponiendo el mecanismo por el cual las hembras elegían, pero no las bases evolutivas que permitieran explicar la evolución de la preferencia de apareamiento en las hembras.
Un mecanismo para la evolución de la preferencia en hembras fue propuesto por Fisher a través del Modelo de escape. Este contempla el beneficio, en términos de éxito reproductivo, que implica para la hembra la preferencia por ciertos machos. Sugirió que un carácter genético nuevo en machos que esté expandiéndose en la población como consecuencia de una determinada ventaja selectiva, puede ser objeto de una cierta preferencia de apareamiento genéticamente determinada en hembras. Así como el carácter beneficioso de los machos se expandirá en la población, el carácter de preferencia, al estar asociado con el beneficioso, se verá expandido también.
El factor de escape al que alude el modelo, el cual permitirá la selección sexual del rasgo. Aquellos machos que posean el carácter preferido por las hembras tendrán cierta ventaja reproductiva porque, al estar expandido el carácter de preferencia, serán más elegidos por las hembras y por lo tanto conseguirán más parejas. Las hembras además de beneficiarse pues su progenie tendrá el carácter beneficioso, incrementará su número de nietos pues sus hijos serán más elegidos.
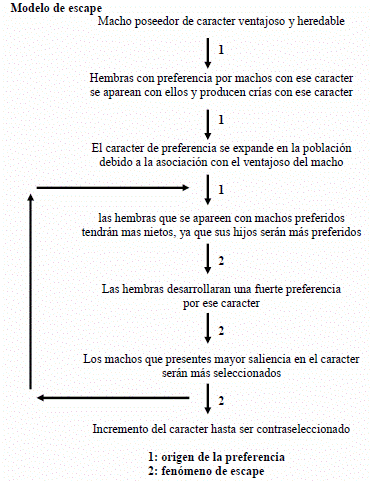
Otras teorías sugieren que la preferencia por un dado carácter neutral en machos puede surgir simplemente por cualquier evento azaroso como la deriva genética y, una vez aparecida la preferencia, se da inicio al proceso de escape.
Puede darse casos donde la hembra no prefiera machos con caracteres neutros, por ejemplo, el de aparearse con individuos de su especie, pero no emparentados, evitando así la hibridización (cruza entre especies distintas) y la endogamia (cruza entre parientes).
SISTEMAS DE APAREAMIENTO
Si un macho por razones filogenéticas se ocupa del cuidado parental no podrá disponer del tiempo y energía necesarios para conseguir nuevas parejas y por lo tanto no practicará esta estrategia. Por otro lado, los factores ecológicos determinan el potencial para monopolizar parejas y por ende limitan la capacidad teórica que ciertos individuos poseerían para acaparar varias parejas.
Si los recursos no son energéticamente defendibles (constreñimientos ecológicos), tendrán que limitarse a conseguir una única pareja y muy probablemente coopere con la hembra en el cuidado parental, ya que deben asegurarse de que la escasa progenie que logren tenga una alta probabilidad de sobrevivir.
La suma de conductas y adaptaciones físicas utilizadas específicamente para conseguir pareja y algunas de las consecuencias sociales de las mismas, constituyen los Sistemas de Apareamiento. Conocer, las estrategias que en teoría permiten a cada sexo incrementar su éxito reproductivo, los rasgos filogenéticos de cada grupo y los constreñimientos ecológicos que modulan el desempeño final, resultará imprescindible para comprender el sistema de apareamiento de una población dada.
Las estrategias potenciales de cada sexo y ciertas variables ecológicas posibilitan explicar la forma que los sistemas adoptan, los cuales están caracterizados por: (i) número de parejas adquiridas, (ii) la forma en que se adquieren las parejas, (iii) el aspecto y características de cada pareja y (iv) el tipo de cuidado parental provisto por cada sexo.
Debido al "efecto Bateman", los animales son potencialmente polígamos. Los experimentos de Bateman estaban caracterizados por trabajar en condiciones ambientales sin limitaciones de recursos o sea sin restricciones ecológicas. Son 2 las condiciones que actualmente favorecen la evolución de la poligamia. En primer lugar, que las parejas o, los recursos para atraerlas sean energéticamente defendibles. El Potencial Ambiental para la Poligamia (PAP) mide cuán económicamente monopolizables son las parejas en determinado ambiente. Depende de varios factores ambientales, pero principalmente del espaciamiento temporal y espacial de los recursos. La segunda condición es la habilidad de los animales para aprovechar un determinado PAP. Esto depende principalmente de factores filogenéticos y de factores ecológicos.
Factores ecológicos claves:
Distribución espacial de los recursos:
Los recursos más críticos son principalmente, el alimento, los refugios, los sitios para colocar huevos o parir, sitios para criar, etc. Existe toda una gama en la distribución espacial de dichos recursos que va desde una distribución uniforme hasta una desigual con áreas de abundancia y otras pobres. El PAP de una distribución uniforme es bajo y la territorialidad con monogamia (al menos aparente) es el sistema resultante. A medida que la irregularidad en la distribución de los recursos aumenta, se incrementa la probabilidad de conseguir parejas adicionales, ya que ciertos individuos podrán acaparar las áreas de altos recursos y con ellas las parejas. En el extremo de la distribución, cuando los recursos están altamente concentrados, el PAP será máximo y un pequeño porcentaje de individuos monopolizará la mayor proporción de los recursos. En este punto la poligamia llegará a su punto máximo, la variación en el éxito reproductivo entre individuos será alto y la selección sexual muy intensa.
Distribución temporal de las parejas :
El recurso cuya variación temporal tiene efectos más marcados en la poligamia es la pareja. Generalmente existe un período limitado durante el cual un individuo es sexualmente receptivo. La sincronización con que los individuos del sexo limitante inician su período receptivo hará que el sexo que compite, también llamado sexo limitado, pueda aparease o no con más de una pareja. Hay un espectro de distribución temporal de parejas potenciales que va desde el sincronismo absoluto hasta asincronismo. A medida que el asincronismo se incrementa, aumentará la posibilidad de que ciertos individuos consigan más de una pareja. Los extremos son perjudiciales. Cuando haya un asincronismo elevado será energéticamente inviable defender durante prolongados períodos los recursos que permitan atraer a las sucesivas parejas. En el extremo opuesto al existir un sincronismo absoluto no hay posibilidad de que se acaparen parejas ya que, aunque ciertos individuos estén capacitados para retener parejas por un tiempo determinado, al ser todas receptivas al unísono y por un tiempo limitado, el individuo monopolizador no podrá copular exitosamente con más que unas pocas.
La Tasa Operacional de Sexos (TOS) define la relación entre el número de hembras receptivas y el número de machos sexualmente activos, para un momento determinado.
Clasificación de los sistemas de apareamiento
Las distintas teorías se basan en dos criterios: el número de parejas y la relación temporal, tanto en la duración como en la simultaneidad de la formación de la pareja.
Tres son los Sistemas de Apareamiento principales: LaPologínia (un macho con más de una hembra), laPoliandría (una hembra con más de un macho) y la Monogamia (un macho y una hembra).
Poliginia.
Ocurre cuando las hembras, por razones ambientales o conductuales, se agrupan y ciertos machos poseen la capacidad de monopolizarlas. Es el sistema que se suele observar en la naturaleza salvo que por factores filogenéticos o ecológicos no se pueda expresar. Tres son las formas de poliginia si se clasifica sobre la base de la forma en que el macho puede acceder y controlar a las hembras , poliginia por defensa de recursos, por defensa de parejas y por dominancia .
Promiscuidad ocurre cuando las hembras son altamente sincrónicas se dan situaciones reproductivas explosivas. Como los machos en sus áreas de despliegue no pueden acaparar parejas debido a la elevada sincronización, se generan apareamientos múltiples durante un período muy corto donde las parejas se forman y deshacen inmediatamente.
Condiciones para la inversión parental por parte del macho:
Generalmente, en la poligamia el cuidado parental es exclusivamente materno. Sin embargo, en ciertos casos es la hembra la que se independiza y el sistema resultante es llamado Poliandria.
Las razones por las cuales el macho puede beneficiarse en criar son: i) Cuando el futuro físico de la pareja es de importancia para el macho. Esto por ejemplo cuando hay fidelidad de temporada en temporada y el macho se beneficia por tener una hembra en buen estado al año siguiente. ii) Cuando la independencia de la hembra implica la posibilidad de que ésta le de nuevas puestas de reemplazo. Las puestas de reemplazo pueden ser de gran importancia en áreas de alta depredación. El principal beneficio para la hembra es la disminución en los gastos metabólicos. La emancipación de la hembra permite una recuperación general luego del costoso proceso de fabricación y puesta de huevos.
Poliandria .
Implica la inversión en el papel de los sexos, esto es, la hembra compite y el macho realiza el cuidado parental. No habría evolucionado si el éxito reproductivo del macho no estuviera limitado por constreñimientos temporales o energéticos y el de las hembras por el acceso a parejas.
Monogamia .
Pareja formada por un macho y una hembra donde ambos están limitados para conseguir parejas adicionales. Las Condiciones para la monogamia son un bajo PAP o un PAP elevado pero baja habilidad para aprovecharlo.
Hay dos hipótesis asociadas con la evolución de la monogamia. Por un lado, “Macho guardián” basada en la idea de que la reducción de los riesgos de paternidad (producto de cópulas extramaritales de su hembra y la competencia espermática) puede lograrse a través del cuidado de la pareja mediante un aumento en la permanencia del macho junto a esta. Otra hipótesis se relaciona con escenarios donde la asistencia del macho en el cuidado parental es decisiva en la supervivencia de la cría.
En los mamíferos gran parte de la inversión parental está asociada a la hembra por razones filogenéticas ya que tanto la gestación como el amamantamiento solo puede ser realizado por estas.
Evolución de las estrategias de elección de parejas en humanos
Según la Psicología Evolucionista la Selección Natural favoreció diseños con un propósito específico debido a que estos pueden resolver mejor los problemas ambientales. Debido a ello, las adaptaciones psicológicas son consideradas con una función de propósito específico y por ende diseñadas para procesar información particular (estímulos o claves ancestrales).
Esto implica entender a la mente humana no como un órgano uniforme multifacético sino como la suma de módulos especializados en problemas particulares. Las adaptaciones psicológicas permitieron maximizar el éxito reproductivo individual en la historia evolutiva del grupo. Por ende, la mente humana al igual que cualquier otro órgano fue diseñado con el propósito de transmitir genes a la siguiente generación.
El concepto de naturaleza humana refiere a que el cerebro humano está compuesto por un gran número de adaptaciones psicológicas que son virtualmente idénticas a lo largo de toda la población. Igualmente se reconoce la existencia de adaptaciones locales y estrategias alternativas, el objetivo principal está centrado en las reglas mentales universales que dan origen, en diferentes condiciones ambientales, a la variación en emociones y comportamientos que han sido documentados a través de las culturas.
Se considera a la Complejidad Adaptativa como cualquier sistema compuesto por varias partes donde los detalles de la estructura de las partes y su ensamble sugieren un diseño para cumplir con alguna función.
Monopolización de parejas, monogamia e infidelidad en humanos:
Existe un acuerdo entre los biólogos evolutivos, de que durante el Pleistoceno las condiciones ecológicas (elevada variación de la probabilidad de conseguir recursos) posibilitaban ocasionalmente la poliginia. La incorporación de la agricultura y la ganadería permitió la acumulación de recursos y un cambio en el arreglo social con un aumento potencial de la poligamia.
Al menos dos razones han sido propuestas. Por un lado, la necesidad de crianza compartida, debido básicamente a la falta de recursos que permitan una crianza independiente por parte de las hembras. Por otro lado, la falta de recursos puede haber promovido que hombre y mujer compartan el alimento y acentuaran su asociación. Una hipótesis adicional supone que la falta de estrógeno en las mujeres (esconder la ovulación) podría ser considerada como una estrategia que permitió a las mujeres asegurar el cuidado por parte del macho celoso e incrementar así la inversión en ellas y sus crías, lo cual llevaría a una monogamia en apariencia. Si el macho desconoce el momento de la ovulación no puede descuidar a su pareja ya que su proximidad y protección será la forma de incrementar la certeza de paternidad, reduciendo los efectos de la competencia espermática.
Si bien distintos estudio muestran cambios en los valores de infidelidad, existe en todos ellos una constante que consiste en una diferencia marcada entre hombres y mujeres, con hombres más infieles que las mujeres en todos los casos.
Tanto el aumento de la infidelidad en mujeres como el aumento del porcentaje del sexo extramarital en hombre pueden ser asociados en parte al denominado efecto Coolidge. Este efecto explica que una de las soluciones psicológicas al problema de conseguir acceso sexual a un mayor número de mujeres está relacionado con la excitación de los hombres por las mujeres. Es decir, el efecto explica como la recuperación de la excitación ante la presentación de nuevas hembras, dando así un mayor impulso para conseguir mantener sexo con varias parejas.
La historia cuenta que el presidente Norteamericano Calvin Coolidge (período 1923-29) y la primera dama (Grace, muy sociable y amistosa) estaban haciendo una visita por separado por unas granjas modelo. Cuando pasaban por el sector de las gallinas y al ver la enérgica cópula que mantenía el gallo con una gallina, la primera dama preguntó sobre la frecuencia con la que el gallo hacia ese trabajo. Docenas de veces por día fue la respuesta del guía. La primera dama solicitó entonces al guía que “le mencione al Sr. presidente este dato”. Cuando el presidente pasó por el mismo sector, el guía cumplió con el encargo de la primera dama y le mencionó al presidente sobre el vigor sexual del gallo, a lo cual el presidente pregunto ¿Siempre con la misma gallina?, Oh no!! replicó el guía, “una diferente cada vez”. “Por favor, hágale saber esto a la primera dama”.
En el caso de los humanos el efecto Coolidge es el responsable del comportamiento denominado “Se le van los ojos” cuando un hombre desvía su mirada para observar a una mujer pasar. Este efecto, en culturas occidentales estudiadas, es el responsable de que la frecuencia de actividad sexual con una pareja decaiga en forma constante con la duración de la pareja, llegando a una 50 % luego de un año de matrimonio y decayendo en forma más gradual posteriormente. Debido a que los efectos son temporo-dependientes, la energía y tiempo invertidos en conseguir nuevas parejas aumenta a medida que la atracción por la pareja formal disminuye, asimismo, el aumento en la inversión del macho en affaire extramaritales va en detrimento del cuidado de la pareja con el resultante aumento de infidelidad en la mujer, explicando así el pico de infidelidad en la mujer.
Un segundo aspecto, relacionado con las estrategias reproductivas del macho, puede contribuir a explicar el pico de infidelidad femenina de mayor edad. Una de las predicciones mejor evaluadas a cerca de estrategias de elección de pareja, está relacionada con la edad de la pareja. Para hombres más que para mujeres la reproducción estará condicionada por la edad de sus parejas. Dos parámetros pueden utilizarse como indicadores de potencial reproductivo. Por un lado, el valor reproductivo, entendido como la descendencia que un individuo es capaz de producir a futuro y, por otro lado, la fertilidad, definida como la probabilidad de reproducción presente (la probabilidad que de una cópula se convierta en un descendiente). En Humanos el Valor Reproductivo de la mujer es máximo en la mitad de la adolescencia (15 años) y luego decrece con la edad. En mujeres la Fertilidad es máxima en los 20 tempranos con valores menores antes y después de esta franja etaria. Debido a que el éxito reproductivo del macho estará condicionado por la capacidad reproductiva de la hembra, la selección a lo largo de miles de generaciones favoreció a aquellos machos capaces de evaluar correctamente la capacidad reproductiva de sus parejas.
Se dispone de Indicadores Temporales para identificar la juventud de una potencial pareja: Apariencia Física: tono muscular, piel suave, cabello brillante, labios carnosos, estructura corporal (90:60:90), simetría facial; y, Apariencia Comportamental: Andar animado, Conductas enérgicas.
Dos predicciones pueden derivarse a partir de este análisis. Predicción 1: Existirá una diferencia en la edad de la pareja elegida dependiendo del sexo, con machos prefiriendo mujeres más jóvenes que ellos. Cuanto más jóvenes, dependerá de sí busca mayor Fertilidad o Valor Reproductivo. Predicción 2: Existirá una diferencia en el atractivo físico, siendo los hombres más exigentes.
Rasgos de atractivo físico como simetría facial, mejillas ruborizadas, labios carnosos y silueta serían los tenidos en cuenta por los hombres al momento de elegir. Estos atributos han mostrado estar relacionados con un mejor funcionamiento del sistema inmune, elevados niveles de estrógenos, estabilidad durante el desarrollo, etc., en general indicadores de un elevado potencial reproductivo.
Devendrá Singh encontró que la relación preferida entre cintura y cadera era “0,7”. La denominada “cintura de avispa” es un rasgo preferido universalmente por los hombres. Mujeres saludables y con un elevado potencial reproductivo poseen una relación cintura-cadera de entre 0,67 y 0,8. Mujeres con una tasa baja mostraron una mayor actividad endocrina en la pubertad. Mujeres en pareja con una tasa alta presentan mayor dificultad para embarazarse y aquellas que se embarazan lo logran a mayor edad comparada con mujeres con menores tasas.
Infidelidad, competencia espermática y celos en humanos:
Debido a que la hembra posee la habilidad de controlar la paternidad de sus hijos a través de las cópulas extramaritales y orgasmos selectivos, es de esperar que, a lo largo de la evolución, se hayan seleccionado positivamente machos con diversas estrategias que les permitieran controlar la paternidad. Los momentos elegidos por mujeres para establecer relaciones extramaritales van en detrimento del éxito reproductivo de la pareja formal. Se encontró que los momentos elegidos por estas para copular en momentos de infidelidad coinciden con los de ovulación .
Baker y Ballis en 1993 identificaron dos fenómenos relacionados con el control de la paternidad por parte de las mujeres. Por un lado, encontraron que el orgasmo femenino que ocurre cercano a la eyaculación masculina resulta en una mayor retención de esperma, con efectos significativos si ocurre entre la eyaculación y hasta 45 minutos posteriores a esta, pero sin efectos de retención de esperma si ocurre incluso un minuto antes. Estos autores mostraron además que en mujeres leales cerca del 55% de los orgasmos fueron de alta retención de esperma (de alta fertilidad). En mujeres infieles solo el 40% de los orgasmos con su pareja formal fueron de alta retención, mientras que el 70% fueron de este tipo con su pareja extramarital. Este enfoque del problema nos permite explicar tanto claves fisiológicas y anatómicas (tamaño testicular, volumen de eyaculación y variaciones en la producción de esperma, etc.) como psicológicas (celos, etc.) asociadas a la infidelidad y al cuidado de la pareja.
Una de las estrategias consiste en modificar la cantidad de esperma puesta en juego con su pareja en función de los riesgos de que esta haya recibido esperma de otros machos. A mayor riesgo mayor número de espermatozoides. Cuanto más tiempo separado pase un hombre de su mujer desde la última cópula, mayor será la cantidad de espermatozoides puestos en juego por este en el siguiente encuentro.
Tanto la producción de esperma como el tamaño relativo de los testículos han mostrado estar relacionados más con el sistema de apareamiento que con el tamaño corporal de la especie.
Entre las claves psicológicas estudiadas relacionadas directamente con los riegos de paternidad y la infidelidad se encuentran los celos y la castidad. Buss evaluó la predicción específica de que los hombres más que las mujeres se preocuparán por la castidad de su futura pareja.
En un estudio se compararon dos eventos estresantes: A) si su pareja mantiene relaciones sexuales con otra pareja o B) si su pareja establece una profunda relación emocional con otra pareja. El 83 % de las mujeres encontraron más preocupante la opción B, mientras que solo un 40 % de los hombres eligieron esa opción. En contraste el 60 % de los hombres experimentaron la relación sexual de su pareja como más estresante mientras que solo el 17 % de las mujeres optaron por la opción A.
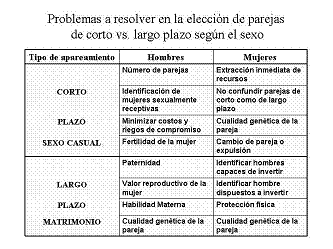
Estrategias Reproductivas y elección de parejas de largo y corto plazo:
Una revisión de las teorías sobre elección de la pareja en humanos permite ver que las decisiones pueden ser clasificadas como “elecciones dirigidas con metas o estratégicas” y “elecciones no dirigidas”.
|
Elección dirigida con metas |
Elección no dirigida |
|
El apareamiento en humanos es estratégico y que hay elección cc o icc, con la intención de maximizar alguna entidad, igualación o balance. · Freud y Jung propusieron que se buscan características que se asemejen al padre de sexo opuesto. · Se busca el “complemento” de “uno”. · Se busca un compañero similar a “uno”. |
No hay estrategia de elección y el mejor predictor de quien se casa con quien es la distancia que los separa. · La elección está más condicionada por la limitante de encuentro que por la búsqueda. · Elementos sociales ponderan la Endogamia (ej.: instituciones educativas sesgan las posibles parejas). |
Las principales limitaciones de estas teorías:
1. Qué motiva a las elecciones. Porqué los humanos preferiríamos similitud en la pareja, semejanza o proximidad. Cuáles serían los orígenes de estas metas.
2. Son modelos simplistas basados en pocos rasgos
3. No pueden generar predicciones en dominios específicos.
4. Asumen que los procesos que motivan a mujeres y hombres son los mismos y por ende no existen predicciones diferenciadas por sexo.
5. Las predicciones son independientes de la duración de la pareja (corto vs. largo plazo).
Gran cantidad de evidencia empírica muestra en particular las diferencias marcadas entre mujeres y hombres con relación a los apareamientos casuales. Comparados con las mujeres los hombres expresan el deseo de tener 4 veces más parejas a lo largo de la vida, experimentan más del doble de fantasías sexuales, muy frecuentemente con cambio de pareja durante un mismo episodio, reducen los requerimientos presentes en las parejas de corto término, están dispuestos a comprometerse en una relación sexual con una pareja pasado muy poco tiempo desde que se conocieron, dedican más tiempo y energía tratando de mantener sexo; el hombre está mucho más predispuesto a tener sexo con un extraño atractivo.
El término de “bajo costo” en relaciones sexuales refiere al bajo costo que implica que los hombres habrán de rechazar a aquellas mujeres que estén buscando un compromiso o una importante inversión antes de tener sexo. A mayor inversión en una pareja, menor será la posibilidad que tendrá el hombre de conseguir parejas adicionales
La naturaleza de la estrategia femenina y sus consecuencias:
La elección de la pareja en las hembras dependerá de los beneficios que pueda obtener de su pareja, los cuales se clasifican dependiendo en primer lugar de si el macho realiza o no, inversión parental. Si el macho no realiza inversión parental las hembras deberán elegir en función de rasgos indicadores de la calidad genética del macho. En especies donde el macho invierte, como es el caso de los humanos, las hembras elegirán de acuerdo a rasgos indicadores de dicha inversión sin descuidar los indicadores de calidad genética. Los indicadores de inversión se clasifican en:
a. Material inmediato para hembra y cría.
b. Incremento en el Éxito Reproductivo futuro de la cría por acceso a beneficios sociales y económicos.
c. Ventajas reproductivas para la hembra y cría si la variación en la calidad del rasgo que permite conseguir los recursos es heredable.
Buss consideró que entre los mejores indicadores de estos tres ítems se encontraba el poder adquisitivo del hombre, y cuando el caballero es joven aun su ambición y laboriosidad. Predijo que las mujeres más que los hombres valorarán estos atributos en sus posibles parejas. Asimismo, Buss observó una elevadacorrelación entre edad del macho y el acceso a recursos, encontrando que en todos los casos las mujeres sistemáticamente prefirieron hombres de mayor edad.
Varios costos se atribuyen a la infidelidad femenina como: pérdida de tiempo en adquirir gametas superfluas, riesgo de transmisión de enfermedades, daños durante la cópula, riesgos de que la pareja engañada abandone la inversión parental en caso de ser descubierta, etc. ¿Cuáles serían entonces los posibles beneficios de la infidelidad femenina? Un posible beneficio está relacionado con la búsqueda del marido ideal. A la mujer le conviene tener un marido con muy buenas cualidades (inteligente, laborioso, con buena salud, corpulento, etc.) y que comparta sus muchos recursos con ella y sus hijos. Pero qué pasa si ambos caracteres no se pueden encontrar en un solo hombre. Solución: conseguir uno que sea bueno y realice la inversión parental (marido) y otro que tenga buenos genes (amante). Esta hipótesis permite entender los resultados arriba enunciados sobre la sincronización de la ovulación con la infidelidad y la elevada frecuencia de orgasmos de alta retención.
Una forma de evaluar los posibles beneficios se obtiene de estudios que analizan las preferencias de mujeres en parejas casuales:
· Que el muchacho gaste mucho dinero en ella desde el principio
· Que le haga un montón de regalos desde el principio
· Que tenga una forma de vida extravagante
· Que sea generoso con sus recursos.
Estos rasgos son muy deseados en amantes, pero no tanto en su pareja estable, ya que según las encuestadas sería un indicador de que puede distraer recursos fácilmente en otras mujeres.
 Un beneficio adicional del sexo casual es poder comparar y establecer si la
elección de pareja está bien hecha. Teniendo en cuenta la importancia de la
elección de pareja en la mujer, este sería un gran beneficio.
Un beneficio adicional del sexo casual es poder comparar y establecer si la
elección de pareja está bien hecha. Teniendo en cuenta la importancia de la
elección de pareja en la mujer, este sería un gran beneficio.
Estrategias reproductivas, problemas adaptativos a resolver y los alcances de la teoría:
Los problemas adaptativos para resolver por los humanos en el intento de buscar parejas dependerán del sexo de quien busca pareja y de si la pareja buscada es de corto o largo plazo.
No todo lo que hacemos nos ayuda a maximizar nuestro éxito reproductivo. Por el contrario, nuestra actividad socializada ha cambiado la forma en que se expresa nuestro comportamiento. Nosotros participamos del erotismo protegiéndonos de la reproducción por métodos anticonceptivos. Pero porque si no queremos tener hijos no abandonamos el sexo. No lo hacemos porque respondemos de acuerdo a una estructura mental que la evolución puso en nuestro cerebro a lo largo de la historia evolutiva del grupo.
Hay una teoría sociológica denominada “debilidad estructural de la mujer” (structural powerlessness) que explica porque las mujeres eligen hombres con mayor poder o recursos. Esta, está basada en la idea de que los hombres excluyen a las mujeres del poder y el acceso a los recursos, los cuales están controlados por hombres. Las mujeres buscarían entonces hombres con poder para acceder ellas al recurso. No es una teoría general, aunque describe bien la generalidad de que los hombres tienen control de los recursos en la mayoría de las culturas. Estas teorías son fenomenológicas donde para cada problema particular se echa mano a un conjunto nuevo de variables que nos permitan resolver el problema.
Semana 8
Modelo Genética del comportamiento. Modelo de los caminos.
El Modelo de los Caminos tiene como objetivo el estudio de la contribución de los factores genéticos y ambientales a las diferencias entre individuos en cuanto a características que hacen a la personalidad y al temperamento.
Las diferencias no son todo el tiempo complejas:
1. Las diferencias en el comportamiento de sistemas complejos pueden ser por razones simples. (Por ej. Un sujeto está más activo que otro porque el primero está lastimado)
2. Es posible atribuir causas a una categoría amplia sin conocer detalles.
Heredabilidad.
Es la contribución de los genes a las diferencias entre individuos de una población, en cuanto a algún rasgo. Es un concepto poblacional, no individual. En sentido amplio refiere a la proporción total de la variación del rasgo debido a los genes, en sentido más acotado refiere a la variación genética trasmitida entre generaciones. Algunos efectos genéticos actúan individualmente, estos se llaman aditivos, debido a que los efectos de genes individuales se suman sobre el rasgo. Hay efectos de genética no aditiva que dependen de la configuración particular de genes presentes: dominancia, depende de la combinación de genes presentes en un locus cromosómico; y, la epistasis, depende la configuración entre loci. Este último afecta sólo una generación ya que no se pasa a descendientes debido a lo que sucede en el reordenamiento de genes en el proceso reproductivo humano. Entonces, la heredabilidad en sentido amplio incluye los efectos aditivos y no aditivos; en sentido reducido sólo incluye los aditivos.
Los rasgos basados en genética aditiva son trasmitibles entre generaciones, mientras que los no aditivos no lo son.
Correlación gen-ambiente.
Ocurre cuando las influencias genéticas y ambientales de un rasgo no son independientes.
Interacción gen-ambiente.
Combinaciones particulares de genes y ambientes pueden conducir a consecuencias no predecibles a partir de ambos factores considerados en forma separada.
El Modelo de los Caminos se aplica para estimar la influencia relativa de genes y ambiente sobre las diferencias individuales en una característica humana. Es decir, se toman las diferencias entre individuos y se determina la influencia de los genes y ambientes sobre el rasgo.
El modelo básico de hermanos Monocigóticos y Dicigóticos:
1- Los hermanos Monocigóticos (gemelos) comparten todos sus genes ya que provienen de la misma cigota formada por las mismas gametas.
2- Los hermanos Dicigóticos (mellizos) comparten entre sí la mitad de sus genes porque provienen de cigotas formadas por gametas diferentes.
De estos surge que la diferencia entre el parecido entre hermanos monocigotas entre sí y el parecido entre hermanos dicigotas entre sí, para un rasgo particular, expresa la mitad del efecto de los genes sobre la variación de ese rasgo.
Para estos estudios se utiliza el coeficiente de correlación que describe el grado en el que dos variables están asociadas, o varían conjuntamente. Dos variables están positivamente correlacionadas si valores altos de una de ellas están asociados con valores altos de la otra. Contrariamente, si valores altos de una de ellas están asociados con bajos valores de la otra, entonces esas dos variables están negativamente correlacionadas. La fuerza de la asociación está indicada por el tamaño del coeficiente de correlación, el cual es un número que varía entre -1.0 y +1.0. Una correlación de 1 (positiva o negativa) indica una asociación perfecta, mientras que una correlación de O significa que no hay asociación lineal entre las dos variables.
El modelo inicialmente asume los supuestos para un rasgo expresado como la extraversión:
1) La correlación gen-ambiente y la interacción gen-ambiente tienen efectos despreciables sobre el rasgo.
2) Los padres de los niños no están correlacionados para ese rasgo. Si los padres estuvieran correlacionados positiva o negativamente, los dicigotas tendrían más o menos, respectivamente, del 50 % de los genes en común.
3) Los efectos de la genética no aditiva son despreciables.
4) El efecto del ambiente sobre ese rasgo es el mismo para monocigotas que para dicigotas.
5) Los gemelos y mellizos son iguales que el resto de la población a la que se quiere generalizar las conclusiones.
Semana 9
Genética del Comportamiento. Teórico.
El comportamiento animal e incluso los rasgos humanos como la personalidad se determinan de forma conjunta por la genética en interacción con el ambiente.
1) Objetivos y Enfoques de la Genética del Comportamiento
El principal punto que aborda la Genética del Comportamiento es el estudio de las bases genéticas de la variación en la conducta entre especies, poblaciones o individuos. Las preguntas planteadas suelen ser:
a. ¿Existen diferencias poblacionales para esta conducta?
b. ¿Las diferencias que se observan son al menos en parte debidas a diferencias genéticas?
c. ¿Existe un patrón de herencia para este rasgo?
d. ¿Cuántos genes controlan este comportamiento?
e. ¿En qué cromosoma está ubicado dicho gen?
f. ¿Cómo afecta la acción del gen la expresión de la conducta?
g. ¿Cuáles son los cambios a nivel del gen o los procesos posteriores hasta la producción de la proteína?
h. ¿Qué proporción de la variación en la conducta alimenticia de estas larvas se debe a factores genéticos y qué proporción a factores ambientales?
i. ¿Qué factores pueden haber actuado haciendo que la selección natural favorezca a uno u otro fenotipo?
Las líneas de investigación se pueden agrupar en dos principales:
· Análisis de los mecanismos genéticos subyacentes a diferencias comportamentales que ocurren en la naturaleza. El objetivo principal consiste en comprender el origen ontogénico y el significado evolutivo de la diferencia comportamental.
· Análisis de los efectos específicos de el o los genes que están influyendo sobre una conducta. Determinar principios generales de como los sistemas (nervioso, endocrino, etc.) se organizan funcionalmente.
Hay dos modelos experimentales predominantes, aunque suelen solaparse en muchos aspectos:
· Analiza aquellos comportamientos que presentan diferencias en condiciones naturales. Las bases genéticas comprometidas en dichos comportamientos presentan efectos directos sobre la conducta. Se utilizan técnicas como la selección artificial para la obtención de líneas puras para cada variante. Se observan directamente las variantes comportamentales en condiciones naturales. Se analizan comparativamente la conducta de gemelos, mellizos, casos de adopción, etc.
· Estudia modelos que presenten alteraciones genéticas del comportamiento, sin importar si estas ocurren o no naturalmente. Se utilizan técnicas destinadas a generar mutantes comportamentales. A través de mutaciones en genes únicos, los patrones comportamentales han sido particionados en sus distintos componentes.
2) Métodos para identificar la relación entre diferencias genéticas y comportamentales
Hay varios métodos utilizados para poner a prueba la hipótesis de que las diferencias genéticas entre individuos son las responsables de las diferencias que éstos presentan en su comportamiento. La hipótesis alternativa está basada en que las diferencias ambientales son las responsables de las distintas variantes comportamentales. Ambas hipótesis parten de la teoría de que, durante el desarrollo de una conducta, la interacción entre el genotipo individual y el ambiente, dan como resultado un fenotipo comportamental particular que dependerá de ambos.
Métodos basados en la comparación de distintos genotipos en ambiente constante:
1. Apareamientos selectivos: se compara y evalúa la influencia de los genes sobre el comportamiento, examinando sistemáticamente el resultado del cruzamiento selectivo de individuos de diferentes especies, poblaciones o distintas líneas dentro de una misma población.
ü Entrecruzamiento entre especies emparentadas: se estudian especies que resultan viables los híbridos estudiando la gerencia de determinados rasgos comportamentales para los cuales dichas especies poseen un desempeño diferente.
ü Entrecruzamiento entre individuos de diferentes poblaciones.
ü Entrecruzamiento entre individuos de diferentes líneas comportamentales dentro de una misma población.
2. Técnicas de selección artificial: se basan en que cierto grado de variabilidad genética existe para dicho rasgo, y que la misma, es al menos en parte, responsable de las diferencias observadas. De ser esto cierto, si durante suficientes generaciones se promueve un apareamiento controlado y selectivo en la población experimental, (solo los portadores de ciertos rasgos se reproducen, y se impide que los individuos con otras características lo hagan), se deberían obtener líneas o linajes puros, cada uno compuesto por individuos con una u otra de las características comportamentales seleccionadas.
3. Estudios comparativos de gemelos: Los gemelos idénticos o monocigóticos, provienen de la duplicación de un único huevo fertilizado por lo que no existe diferencia genética alguna entre ellos. Los mellizos o gemelos dicigóticos provienen de dos óvulos distintos fertilizados por dos espermatozoides distintos, por lo que son genéticamente equivalentes a dos hermanos completos (hijos del mismo padre y madre). Asimismo, tanto gemelos entre sí, como mellizos entre sí, estuvieron sometidos a ambientes muy semejantes y donde la gran diferencia entre unos y otros está casi exclusivamente basada en diferencias genéticas (los primeros son idénticos y los segundos comparten en promedio un 50% de sus genes).
Cuando gemelos monocigóticos (GM) son comparados entre sí en cuanto a una serie
de rasgos psicológicos, el grado se semejanza puede ser estimado mediante
correlaciones. Los gemelos monocigóticos muestran sistemáticamente una mayor similitud que los dicigóticos, lo cual, debido a que ambos tipos se desarrollan bajo ambientes similares, implicaría que diferencias genéticas son las responsables de distinguir a los "GD" o de hacer más semejantes a los "GM". Sin embargo, al desconocerse el verdadero efecto del ambiente sobre varios de los rasgos psicológicos muchos psicólogos cuestionan la relevancia de dichas comparaciones. Estos basan sus críticas en la idea de que, los "GM" con seguridad experimentan un ambiente más semejante que los "GD". Por lo tanto, si el efecto del ambiente fuese importante, esta diferencia (incluso en el caso de ser menor), podría ser radical al momento de explicar las diferencias observadas, o más aún, justificar porque los "GM" son tan parecidos.
Métodos basados en la respuesta a ambientes alternativos:
1. Cambios ambientales y territorialidad en arañas: La araña del desierto (Agelenopsis aperta) construye tela y defiende un área alrededor de la misma. Esta conducta territorial impide que otras arañas construyan telas cerca de la propia. Entre otros, factores climáticos y de abundancia de presas, son los responsables del cambio en el tamaño territorial.
Las diferencias poblacionales en el tamaño territorial podrían deberse a diferencias genéticas por lo cual por herencia se mantendría el carácter. Otra hipótesis considera probable que los individuos evalúen la oferta local de alimento o factibilidad de caza y ajusten el tamaño territorial a esta variable.
Riechert , realizó una serie de experiencias para poner a prueba estas dos hipótesis. Obtuvo muestras de dos poblaciones: una ribereña, de territorios pequeños y una de pastizal de territorios grandes. 3.8m2). Las arañas fueron llevadas al laboratorio y expuestas a un régimen de densidad de presas constante para ambas y equivalente al promedio de los valores encontrados en todos los ambientes naturales. Una vez aclimatadas al laboratorio, cada araña fue colocada individualmente en un cilindro metálico que impedía detectar la presencia de otras arañas. Como resultado de esto, hubo un aumento de la proximidad entre las telas de diferentes individuos. Posteriormente, el cilindro fue removido y la conducta territorial de las arañas analizada. Las arañas ribereñas establecieron sus pequeños territorios, y las de los pastizales sus territorios de mayor extensión, sin que el cambio en las condiciones de oferta de presas alterara sus conductas territoriales. El restablecimiento de los territorios con magnitudes semejantes a las naturales hizo que ciertos sujetos no consiguieran espacio suficiente, lo cual no afectó el tamaño territorial, sino que por el contrarios dejó sin territorio a varios animales. Estos resultados sugieren que las diferencias poblacionales se deben a diferencias genéticas o que al menos ni la densidad de presas ni la de conespecíficos condicionan el tamaño territorial .
2. Adopción y crianza diferencial de individuos emparentados: En aquellos casos en que las crías puedan ser transferidas de los padres originales a padres alternativos, de la misma o distinta población, raza o especie, se estará en condiciones de comparar individuos genéticamente similares (esto dependerá del grado de parentesco) criados en ambientes diferentes.
a. Adopción Interespecífica.
b. Adopción Intraespecífica en humanos: Estudios complementarios de adopción, intentaron evaluar la influencia ambiental sobre determinados rasgos cognitivos. El ambiente compartido, entre hijos, es considerado como componente de la varianza ambiental que afecta las diferencias individuales en tests de Inteligencia (IQ). En 11 estudios, la correlación promedio de los valores de IQ entre hermanos adoptados (no biológicos) fue de 0,3. Esto sugiere que el 30% de la varianza en los valores de IQ es debida a haber compartido las influencias ambientales
Estudio de las variaciones comportamentales cuantitativas.
Las características de los sujetos son predecibles por lo que se simplifica la contabilización de los distintos fenotipos y se pueden realizar análisis estadísticos confiables y robustos.
Muchos de los caracteres comportamentales son de variación continua y las poblaciones no pueden ser clasificadas en categorías discretas entre las cuales no existan intermedios. La diferencia entre ambos tipos de caracteres radica en que en el primer caso uno o muy pocos genes son los responsables de actuar en el desarrollo de la conducta. En el segundo, la variación continua puede ser resultado de la acción de un número variable de genes actuando independientemente sobre el rasgo en cuestión, sin poder descartar que factores ambientales o la combinación de ambos sean los responsables.
Estos caracteres de variación continua son denominados usualmente caracteres cuantitativos y el estudio de la variación genética comprometida es llamada "Genética Cuantitativa".
1. Grado de determinación genética y heredabilidad:
El fenotipo (F) de un individuo está determinado por factores ambientales (A) y genéticos (G) los cuales se combinan en forma aditiva. Si VF representa la variación fenotípica total observada en una población, VG: el componente genotípico de la variación total y VA: el componente ambiental, la relación entre dichos factores será: VF=VG+VA+VI; donde VI: representa la variación atribuible a la interacción entre genotipo y ambiente.
La heredabilidad expresa la proporción del variación fenotípica total que es atribuible al componente genético. Es un concepto poblacional, que suele utilizarse. En un sentido amplio se refiere a la proporción de variación que es debida a los genes y en un sentido estricto, se refiere sólo a aquella parte de la variación genética que se transmite a través de las generaciones. Esta diferencia está basada en que para algunos efectos genéticos los genes actúan individualmente, y son denominados "aditivos", ya que los efectos individuales se suman unos a otros en un efecto general. Existen efectos denominados "no aditivos" que son el resultado de efectos genéticos de dominancia y epistasis, los cuales dependen de la configuración particular de genes que están presentes. Los efectos "no aditivos" contribuyen a la variación genética en una población dada en cada generación, pero debido a la predisposición de los genes durante la reproducción no se transmiten de padres a hijos.
La heredabilidad (h2) en el sentido amplio incluye ambos tipos de efectos y es también denominada "grado de determinación genética" (DG), calculándose como: h2=DG=VG/VF=VG/VG+VA (considerando VI=0). El rango de valores de h2 es: 0<=h2<=1.
Una forma de evaluar el grado de determinación genética consiste en estudiar diferentes cruzamientos entre líneas endocriadas para cierto carácter conductual.
La heredabilidad no es una medida invariable de la influencia de los genes en un carácter particular, sino un indicador de la manera en que los efectos genéticos alteran su expresión en un ambiente particular, para una población determinada, en un momento dado.
Las estimaciones de heredabilidad pueden estar sujetas a errores de diferentes fuentes, siendo las más importantes la interacción gen-ambiente y la correlación gen-ambiente. La interacción gen-ambiente se refiere al hecho de que una particular combinación de genes y ambiente genera consecuencias no predecibles al considerar a ambos por separado .
La correlación gen-ambiente se refiere al hecho de que no existe independencia entre las influencias genéticas y ambientales sobre un rasgo comportamental. Este tipo de correlación es denominada "pasiva" ya que ocurre independientemente del comportamiento.
Métodos de alteración del genoma como generador de variantes conductuales:
1. Mutaciones.
Gran parte del conocimiento acumulado sobre el efecto específico de los genes sobre el comportamiento ha partido del estudios sistemático de mutantes genéticos.
Muchas mutaciones surgen de forma espontánea en la naturaleza. Los mutantes comportamentales poseen alteraciones en diferentes aspectos, con la ventaja de que la mutación comportamental está acompañada en muchos casos, de alteraciones en rasgos que permiten fácilmente identificar la variante. Así, por ejemplo, un cambio en la coloración general del animal es indicador de ciertas modificaciones en la cópula.
2. Mutantes, cambios en el sistema nervioso y diferencias comportamentales:
a. Alteraciones en la bioquímica de sistema nervioso: el sistema nervioso es el sistema que mayor efecto posee sobre la conducta de la mayoría de los animales. Cada célula nerviosa está dotada de un conjunto de genes que se encargará a lo largo de su desarrollo de controlar todas sus funciones, entre ellas la química de la célula. Así, la química de la célula nerviosa condiciona la transmisión de los mensajes a través de esta, y de una en otra. Una pequeña alteración en un gen puede ocasionar cambios neuroquímicos de efectos considerables en la conducta. Este es el caso de alteraciones en neurotransmisores y ciertos comportamientos complejos en humanos.
La dopamina, serotonina y noradrenalina serían las responsables de intermediar, en muchos caso mediante mecanismos no conocidos, en diferentes conductas complejas en humanos. Modificaciones en el dosaje de estos neurotransmisores ha mostrado relacionarse con alteraciones comportamentales como: esquizofrenia, retardos mentales, agresión, etc. Las evidencias de que la dopamina está relacionada con la esquizofrenia resultan de los efectos de drogas antipsicóticas llamadas "neurolépticos". Estas, actúan bloqueando los receptores para dopamina y han mostrado buenos resultados en el tratamiento de la esquizofrenia. La enzima L-dopa, por el contrario, incrementa los niveles de dopamina haciendo que pacientes con Parkinson (deficiencia de dopamina) muestren cuadros psicóticos con rasgos semejantes a una esquizofrenia.
En ciertos casos los cambios observados están relacionados con los dosajes del neurotransmisor debido a cambios en la producción de enzimas responsables del metabolismo de los mismos, como el caso de las Monoaminoxidasas (MAO) A y B. Familias con bajos niveles de MAO tienen 8 veces más intentos de suicidio que aquellas con altos niveles de MAO.
b. Alteraciones estructurales del sistema nervioso: el modelo de nematodes (Caenorhabditis elegans) es el utilizado para el análisis de relaciones específicas entre mutaciones, variantes estructurales del sistema nervioso y comportamientos alternativos. Las ventajas del modelo consisten en que un es un organismo simple del cual se conoce profundamente su neuroanatomía y neurofisiología, y el cual posee un considerable repertorio de comportamientos. Sumado a esto, alrededor de 250 genes han sido identificados como reguladores de su conducta mediante técnicas de mutación. El conocimiento detallado de este organismo ha llegado a tal extremo que tanto el número de neuronas (302), sinapsis (5000) y uniones neuromusculares (2000) como la posición exacta de las mismas, están al alcance de cualquier investigador que desee analizar su comportamiento. Además, mediante técnicas láser se ha logrado identificar el rol específico para 40 de las 118 clases estructurales de neuronas. En cuanto a la bioquímica, la gran mayoría de los neurotransmisores y receptores son familiares para cualquier neurobiólogo.
Se conocen, para esta especie, varias formas de aprendizaje no asociativo. Diferentes taxias, en especial con respecto a olores hacia los cuales responde incluso en gradientes de variaciones graduales y para unas 60 compuestos volátiles. Posee respuestas sensoriales a feromonas, etc.
Estudios detallados se han hecho con respecto a la respuesta a la estimulación mecánica. Existen diferentes mutantes para la sensibilidad táctil, los cuales son muy específicos en sus efectos. Estos mutantes responden y se mueven normalmente pero solo ante estimulaciones táctiles fuertes. Los receptores al tacto son un conjunto de 6 neuronas mecanoreceptoras. Los mutantes posibles para este rasgo, están ubicados en 12 loci diferentes y sus efectos van desde la pérdida de microtúbulos citoplasmáticos, pasando por alteraciones en su contacto con a hipodermis, hasta la desaparición casi completa del mecanoreceptor.
c. Mutantes, propiedades eléctricas de la membrana celular y comportamiento : Los paramecio contacta con un obstáculo e invierten la dirección de su marcha para luego reiniciarla con un cambio en la orientación. Es una respuesta de evitación normal que depende de una secuencia específica de cambios químicos y comportamiento resultantes: 1) estímulo táctil 2) cambio en la permeabilidad de membrana 3) entrada de iones (calcio) Ca++ 4) Cambio positivo en el potencial de membrana 5) cambio en la actividad de las cilias 6) cambio en la dirección de desplazamiento 7) sistema de bombas sacan Ca++ hasta restablecer el potencial de membrana original (aproximadamente 2 segundos) 8) reinicio de la dirección de natación original.
Genética del Comportamiento y Desarrollo:
Cada comportamiento es el resultado de la interacción entre estímulos externos y condiciones internas. Como los sistemas internos de procesamiento de información son establecidos y van siendo funcionales durante el curso del desarrollo, cualquier conducta está limitada por la forma en que la información es procesada por cada sujeto, a lo largo de la distintas etapas ontogénicas. Entre las acciones más notorias de los diferentes genes que contribuyen a rasgos conductuales, está el de los genes reguladores que actúan coordinando los complejos procesos embriológicos. El producto de la actividad de dichos genes está condicionado por el momento particular en el desarrollo y las condiciones ambientales al momento, en que el gen comienza a expresarse.
El conocimiento del desarrollo del comportamiento depende de la identificación de los factores que pueden influenciar dicho proceso y la forma en que lo hacen. Es esencial la información sobre el genoma ya que es la mayor fuente de información interna para el sistema en desarrollo.
Controversia Nature (control genético) - Nurture (influencias ambientales).
1. Diferencias Genéticas, Índice de Inteligencia (IQ) y la controversia Nature-Nurture:
Para gemelos idénticos criados juntos la correlación de IQ oscila entre 0,5 y 0,9. Bouchard y colaboradores, estudiando mellizos y gemelos, estimaron que la heredabilidad del IQ es de 0,7. Debido a que la heredabilidad de los escores de IQ es generalmente significativa, suele interpretarse erróneamente que la "Inteligencia" está genéticamente determinada. Los resultados encontrados al comparar con las técnicas enumeradas solo permiten decir: Diferencias genéticas entre las personas estudiadas hasta la fecha contribuyen en forma significativa en las diferencias en escores determinados en tests de IQ. Esto no implica que el IQ sea "genético" o "genéticamente determinado" o "heredado" en el sentido de no necesitar de influencias ambientales para su desarrollo.
Se diferencian dos conceptos:
a) Reconocer que existen influencias genéticas importantes en la generación de las conductas, así, diferencias genéticas acompañan a diferencias en el rasgo, incluso en comportamientos complejos.
b) Cuantificar la independencia con respecto al ambiente por parte de determinadas conductas. Esto es para decir que una conducta es innata descartando el efecto ambiental.
2. Ontogenia del canto en aves y la controversia Nature-Nurture:
Un ejemplo es la ontogenia de la conducta de canto en aves. Hay modelos que buscan representar los estadios y los efectos de genes y ambiente en el desarrollo de esta conducta. El modelo del Templado auditivo inspiró muchos diseños experimentales los cuales están estructurados administrando la experiencia ambiental a la que están sometidas las aves durante su desarrollo. Ciertos diseños eliminan la retroalimentación auditiva, otros solo la posibilidad de que el pájaro pueda escuchar a otras aves, etc. En todos los casos se intentan dos manipulaciones alternativas: un mismo genotipo en diferentes ambientes o distintos genotipos en un único ambiente.
Por ejemplo, los distintos sujetos de una camada son distribuidos en tres posibles grupos experimentales donde cambian las condiciones ambientales, a) los sujetos son aislados de conespecíficos, b) los sujetos son ensordecidos y c) los sujetos son entrenados con grabaciones de cantos conespecíficos. Dependiendo de las características del canto final que produzcan se concluirá que:
1) Si todos cantan parecido independientemente del tratamiento entonces se dirá que el canto es instintivo o puramente genético.
2) Si "a" y "c" cantan normalmente, pero "b" no, se dirá que el canto solo depende de poder escucharse durante la ontogenia sin que el uso de modelos afecte el producto. Por lo tanto, el canto tiene un componente ambiental pero regido por un modelo innato.
Semana 10
Origen del lenguaje. Práctico.
Los estudios sobre adquisición de un lenguaje en simios como una herramienta para rastrear el origen del lenguaje humano.
Visiones del lenguaje.
Ciertos autores consideran que el lenguaje debe ser visto como un emergente comportamental y como un sistema cognitivo que debe ser estudiado dentro de un marco comparativo y de desarrollo, por lo que la adquisición del lenguaje debe estudiarse también en no-humanos. Chomsky y Skinner fueron uno de los propulsores de las perspectivas nativistas y empiristas, del origen del lenguaje.
Nativistas (Chomsky)
- Capacidad única de nuestra especie.
· Existencia de “universales” gramaticales que trascienden los sistemas específicos de lenguaje.
- Lo innato es la capacidad de hablar cualquier lenguaje.
· Los niños adquieren un dominio de reglas gramaticales complejas al escuchar a otro hablar.
- Énfasis en la producción del habla.
Empiristas (Skinner)
· Sistema complejo de comportamiento que puede ser adquirido a través de procesos generales de aprendizaje que no son específicos de nuestra especie.
· Aprendemos a hablar como aprendemos cualquier cosa.
· Otros medios además del habla pueden servir al lenguaje y sus funciones.
· Lenguaje como emergente comportamental que debe ser estudiado en un marco compartido y de desarrollo.
- Debe estudiarse en animales no humanos.
- Importancia de la comprensión como fundamento del lenguaje.
Definición del lenguaje.
Desde perspectiva de trabajo, Rumbaugh y Savage-Rumbaugh dicen que, el lenguaje, es un sistema multidimensional, neuro-comportamental, que provee la construcción y el uso de símbolos, de una manera que facilita el envío y la recepción de información y de ideas nuevas entre individuos. Los significados de los símbolos en este sistema están básicamente definidos y modulados a través de interacciones sociales.
En esta perspectiva sería considerado como un sistema abierto a la incorporación de nuevos símbolos, nuevas funciones y usos, y a la atribución de significados a través de usos sociales novedosos de los símbolos.
El habla representa un sistema que, junto al oído permite una rápida expresión del lenguaje. No es el lenguaje.
EL SENTIDO DE LOS ESTUDIOS SOBRE LENGUAJE EN PRIMATES: RECONSTRUCCIÓN DE NUESTRO PASADO HOMÍNIDO.
Ubicación evolutiva del hombre.
De acuerdo a la clasificación utilizada por los biólogos, el hombre ocupa la siguiente posición:
- Reino: Animalia
- Phylum: Chordata
- Clase: Mammalia
- Orden: Primate
- Familia: Hominidae
- Género: Homo
- Especie: Homo sapiens
Cada especie hereda un juego de atributos anatómicos y comportamentales que caracterizan al orden como un todo, pero también cada especie es única, el resultado de su propia historia evolutiva. Los primates modernos pueden ser divididos en cuatro grupos:
1. Prosimios (lémures y társidos)
2. Monos del Nuevo Mundo (monos tití, monos araña, monos aulladores, etc.)
3. Monos del Viejo Mundo (macacos, babuinos, colobos, etc.)
4. Hominoides (simios y humanos)
Los monos y simios se los suele conocer como antropoides.
Homínidos primitivos
En la familia Hominidae se encuentras los homínidos primitivos. Los primeros surgieron entre los 3,75 y los 4 millones de años atrás en Tanzania y Etiopía. Estos serían los Australopithecus.
Hace aproximadamente dos millones y medio de años, los homínidos experimentaron una radiación adaptativa. En la parte oriental del continente una especie de Australopithecus, A. boisei, convivía con un homínido de encéfalo desarrollado, hoy catalogado como Homo habilis, con el que empieza el linaje Homo. Pero hace alrededor de 1,75 millones de años, H. Habilis desapareció de África, siendo sustituido por un homínido provisto de un encéfalo mayor: Homo erectus. Apareció en África hace 1.8 millones de años, y hace 1 millón de años vivía en el sudeste y este de Asia, permaneciendo en esa área por lo menos hasta hace 300.000 años.
Entre 400 y 300 mil años atrás aparecen los primeros Homo sapiens arcaicos, que dan lugar a dos líneas: por un lado, los neanderthales (Homo sapiens neanderhalensis); y por otro, a los humanos modernos (Homo sapiens sapiens). Los humanos modernos más antiguos conocidos están datados en unos 100.000 años.
Reconstrucción de la forma de vida homínida
La forma de vida de los primeros homínidos puede mostrar los procesos por los que nosotros evolucionamos y los mayores eventos que marcaron el origen de la humanidad.
Para esto se utilizan evidencias a partir de los registros fósiles y arqueológicos y un marco para interpretarlas y hacer inferencias a partir de las mismas, que surge de observaciones y experimentos realizados en la actualidad.
Por ej.: La biomecánica y la anatomía funcional permiten hacer inferencias sobre los patrones de movimiento a partir de la morfología de los huesos fósiles.
LOS ORÍGENES DEL LENGUAJE
Evidencias fósiles
Los humanos sobresalen en tres aspectos principales: su esqueleto está adaptado para la locomoción bípeda, tienen mandíbulas y dientes modificados, y, sobre todo, tienen cerebros muy grandes. Estas tres características pueden estar íntimamente conectadas: la locomoción es crucial para la obtención de alimentos, los dientes y las mandíbulas para el procesado del alimento; la energía obtenida del alimento permite afrontar los altísimos costos energéticos del cerebro.
Entonces, la evolución del cerebro humano puede explicarse en dos niveles:
1. El incremento en el tamaño desde los australopithecinos hasta los primeros Homo probablemente dependió de un cambio en los hábitats de alimentación, que condujo a un incremento en el recambio energético.
2. Las presiones selectivas específicas pueden haber conducido a cambios en las conexiones internas del tejido neuronal en expansión, que permitieron el aumento en la capacidad de resolución de problemas, el origen de la fabricación de herramientas complejas y, la emergencia del lenguaje y de la cultura.
El cerebro de los primates.
Hay un consenso sobre que el primate promedio tiene un cerebro más grande en relación a su cuerpo que muchos otros mamíferos. Aunque no se puede terminar de comprender esto. Asimismo, las regiones de áreas corticales y medulares asociadas al control del tacto y movimientos en los primates son particularmente fuertes. Aunque los músculos de la boca y la garganta están bien representados en la corteza motora primate, la producción de sonidos vocales no está bajo control motor. Las vocalizaciones dependen mucho más de la corteza límbica y de numerosos sistemas subcorticales.
En cuanto a las capacidades auditivas, en monos la percepción de secuencias rápidas de sonidos se realiza mejor en el lado izquierdo del cerebro, y en macacos japoneses el análisis de las llamadas vocales entre miembros de la especie es llevado a cabo principalmente también en el «hemisferio izquierdo. Hay aquí un paralelismo con la lateralización de los procesos del lenguaje en el cerebro humano.
El cerebro humano.
El tamaño del cerebro se lo relaciona al uso de correcciones alométricas para corregir el efecto de las diferencias entre las tasas de crecimiento del cerebro y el cuerpo dando por resultado un coeficiente de encefalización.
El estudio de los cráneos fósiles encontrados y de sus endocastos (impresiones sobre el cráneo de la superficie del cerebro) permitieron establecer una tendencia hacia una encefalización creciente a lo largo de la evolución humana. La encefalización habría superado verdaderamente el rango de los simios haría unos 2 millones de años, con la aparición del primer miembro reconocido de nuestro género, Homo habilis, con un cerebro de aproximadamente 750 grs. Se cree que los cerebros de los primeros Homo sapiens y los neandertales tendrían un tamaño similar al nuestro.
Debido a que el cuerpo de las mujeres no sería capaces de sostener el nivel de crecimiento fetal y su canal de parto resultaría incompatible con una locomoción bípeda, las crías humanas nacen neurológicamente inmaduras, completando su desarrollo postparto.
Áreas cerebrales del lenguaje.
No existe aún una teoría comprehensiva de cómo se produce el lenguaje.
El habla humana depende de las regiones de la corteza motora que controlan la boca, la lengua y la laringe, y de áreas en frente de esta. Las áreas cerebrales especializadas se encuentran en la región que rodea a la fisura de Silvio en el hemisferio izquierdo. Este también contiene centros para la percepción auditiva, la percepción táctil, y el control motor de la cara, la boca y la laringe.
Los estudios de daños cerebrales y de electroestimulación implicaron a lasáreas de Broca y de Wernicke en la producción y comprensión del lenguaje. Se demostró que diferentes aspectos del lenguaje como la secuencia de movimientos orales, la identificación de fonemas, la gramática, la lectura, y la memoria verbal de corto plazo, se encuentran controladas por diferentes subdivisiones de esas áreas y también por otras áreas diferentes a las clásicas. Además, la distribución de las funciones del lenguaje varía de persona a persona.
Una de las mayores especializaciones del cerebro para el lenguaje es el papel diferencial de los dos hemisferios. Aunque aún no se conoce la causa de esta lateralización.
En el 90% de toda la gente el hemisferio izquierdo es el dominante en la producción y el análisis del lenguaje. Esto incluye alrededor del 95% de todos los diestros y el 70% de todos los zurdos. La tendencia que la mano dominante acompañe a la representación del lenguaje en el hemisferio izquierdo sugiere que la habilidad manual y la habilidad para el lenguaje están ligadas.
Con el objetivo de inferir las capacidades lingüísticas de nuestros ancestros se estudiaron los endocastos. La cisura de Silvio se ha encontrado en algunos endocastos de miembros arcaicos del género Homo. En humanos modernos ésta está correlacionada con el gran tamaño del área de Wernicke en el hemisferio izquierdo. En un comienzo se creía que esto indicaba que había presencia de un lenguaje hablado, pero estas asimetrías fueron encontradas en monos y simios modernos, que no hablan. Sin embargo, las asimetrías corticales en el registro fósil nos pueden decir algo acerca de los orígenes de asimetrías funcionales más generales.
Se ha demostrado la presencia de los surcos asociados al área de Broca en Homo habilis y H. erectus, marcas que no aparecen en los endocastos de los Australopithecus. Esto indica que el área de Broca apareció tempranamente en los primeros Homo, si se la define en términos morfológicos; pero no implica que el lenguaje haya aparecido en ese momento. En primer lugar, hay una gran variabilidad en la presencia y posición de esos surcos en el cerebro humano, y cierta inconsistencia en su asociación con las funciones del lenguaje. Y, además, se ha demostrado la existencia de una zona homóloga en el cerebro de monos, de manera tal que su presencia en los fósiles no indica la aparición de una estructura totalmente nueva.
Reorganización del cerebro
Las evidencias provenientes del estudio comparado de la estructura cerebral sugieren que la base de las diferencias específicas en la estructura del cerebro no es la adición de nuevas estructuras, ni el cambio en las conexiones de estructuras ya existentes. En cambio, dichas evidencias apuntan a la reorganización sistemática, la elaboración o reducción de estructuras, o cambios en la proporción de conexiones existentes.
El reclutamiento de circuitos y estructuras cerebrales existentes estaría plasmado en la evolución del lenguaje humano. El lenguaje no podría haber sido el resultado de la adición de nuevas estructuras porque está controlado no por una única estructura cerebral, sino por una red de áreas corticales interdependientes, cada una de las cuales colabora en una función particular. Estas estructuras tienen su contraparte en los cerebros de primates (incapaces de hablar).
La evolución ha "reclutado", para propósitos que tienen que ver con el lenguaje, estructuras que cumplen otras funciones en primates no-humanos. En este proceso de reclutamiento, dichas áreas se modificaron para adaptarse a requisitos funcionales diferentes y mucho más exigentes. El tamaño de distintas estructuras cerebrales sugiere que el agrandamiento del cerebro durante la evolución humana no fue uniforme para todas las áreas, y siguió tendencias diferentes a la expansión cerebral de otros primates. Los cambios en el tamaño relativo de estas áreas también cambiaron su función alterando conexiones entre estructuras.
El habla humana tiene características particulares que plantean requisitos específicos. La habilidad para combinar un gran número de sonidos para formar unidades más largas, palabras y frases, hace posible la complejidad sintáctica del habla. La sintaxis y la gramática le implican al cerebro procesos complicados y exigentes.
El aparato fonador.
El lenguaje humano involucra mecanismos innatos, anatómicos y neurales, trasmitidos genéticamente. Particularmente los relacionados al habla. Son características únicas en los humanos indicando una evolución para aumentar la capacidad de usar el lenguaje.
El habla nos permite transmitir segmentos fonéticos (que se ajustan, aproximadamente, a las letras del alfabeto) a una tasa considerable de más de 25 por segundo. En contraste, es imposible identificar información no hablada a una tasa mayor a 7 o 9 ítems por segundo.
La alta tasa de transmisión del habla sería fundamental en la habilidad lingüística ya que permite que ideas complejas puedan ser transmitidas dentro de los límites de memoria a corto plazo.
Esta alta tasa de transmisión se logra a través de la generación de patrones de frecuencia formante y de la rápida producción de señales por el específico tracto supralaríngeo humano y sus mecanismos de control.
Hasta los tres meses de edad, los bebés humanos tienen un tracto aéreo similar al del resto de los mamíferos, tal que pueden respirar y beber al mismo tiempo. En los humanos adultos, en cambio, la laringe ha descendido adentro del cuello. La lengua forma tanto el límite anterior de la faringe como el límite inferior de la cavidad oral.
Durante el desarrollo, el paladar humano se mueve hacia atrás a lo largo de la base del cráneo, el cual se reestructura para adquirir el tracto humano supralaríngeo. Los humanos son menos eficientes para masticar, porque el lecho de la boca y la mandíbula inferior se han reducido en comparación con primates no humanos y con los homínidos arcaicos. Además, esta reducción del paladar y de la mandíbula hace que nuestros dientes se apretujen, lo que puede aumentar las probabilidades de contraer una infección.
Estas deficiencias asociadas al habla humana se ven compensadas por el incremento en el rango fonético del tracto supralaríngeo, el cual puede cerrar el camino que conduce al aire desde la nariz hacia la boca, pudiendo así producir sonidos no-nasales.
Habilidad manual y fabricación de herramientas
La fabricación y el uso de herramienta brindan información sobre las capacidades cognitivas de nuestros antepasados, así como sobre su cultura y forma de transmitirla.
Las primeras evidencias para las habilidades manipulativas humanas provienen de los huesos de manos y brazos de los australopithecinos. Sus manos tienen una mezcla interesante de características humanas y de otros primates como el chimpancé. Ellas muestran un patrón anatómico primariamente adaptado para la locomoción, con sólo ligeras modificaciones para la manipulación. No hay evidencia que los australopithecinos hicieran y usaran más herramientas que los chimpancés modernos.
Los primeros progresos manipulativos complejos aparecen en el género Homo, en las especies H. habilis. Los huesos fosilizados de sus manos indican un patrón humano moderno de movilidad del pulgar y se mantiene la poderosa musculatura de flexión de los dedos. Esta especie muestra la mayor expansión y, probablemente, la mayor reorganización del cerebro, una bipedestación totalmente terrestre, y la fabricación y uso de herramientas de piedra.
El lenguaje está asociado a la lateralización del cerebro. Las herramientas solían ser utilizadas en la mano derecha, había una preferencia. Lo mismo sucede con los primates no humanos.
PRINCIPALES ESTUDIOS SOBRE APRENDIZAJE DE UN LENGUAJE EN SIMIOS
El hombre y el chimpancé son los que se encuentran genéticamente más relacionados, comparten el 99% del material genético. Es por esto que las primeras investigaciones que abordaron el tema del lenguaje y su adquisición tomaron a los chimpancés como material de estudio.
Proyecto Vicky
En los 50 se pretendió enseñarle a hablar a los simios. Hayes y Hayes se destacaron con una chimpancé llamada Vicky, la cual fue criada como un niño y con un entrenamiento formal. Este entrenamiento consistía en sesiones en la que le mostraban distintos objetos mientras pronunciaban el nombre de cada uno. Pero el resultado no fue satisfactorio: Vicky produjo sólo 4 palabras apenas inteligibles.
En el momento no se sabían las limitaciones anatómicas y neurales en el tracto vocal de los monos impedía la producción de sonidos necesaria para el habla humana y, por lo tanto, de estos trabajos se infirió que eran incapaces de adquirir lenguaje.
Sin embargo, la limitación implica el lenguaje oral. Surgió una necesidad de una reconcepción del lenguaje, en donde se consideró al habla como una de las formas que éste puede adoptar.
Proyecto Washoe
En la década del '60, Gardner y Gardner estudiaron la posibilidad de adquisición del lenguaje de señas de sordo-mudos. El entrenamiento del chimpancé fue llevado a cabo modelando sus manos en la posición adecuada para cada símbolo gestual y recompensándola cada vez que producía una seña acertada. Luego de un período de aprendizaje, Washoe fue capaz de usar un gran número de señas en forma individual y encadenada, incluso en test ciegos -en ausencia de entrenadores- y en forma solitaria. Si bien los investigadores estaban de acuerdo en su capacidad gestual -es decir, su habilidad para referirse a un objeto mediante una seña-, quedaban dudas de si estas señas eran utilizadas realmente como nombres o eran trucos para conseguir lo que pedía. Este tipo de aprendizaje es llamado discriminación condicionada, un término que significa que un animal puede aprender distintos sets de respuestas y ejecutarlos independientemente frente a distintas situaciones.
Los Gardner sostenían que Washoe era capaz de usar señas y no estaba condicionado justificándolo con las pruebas de vocabulario.
Frente a estos resultados, varios científicos abocados a la postura nativista hicieron hincapié en que la esencia del lenguaje humano no es el uso de palabras individuales sino la combinación de éstas siguiendo reglas específicas y que es la utilización de estas reglas lo que es propio de la especie humana.
Proyectos Sarah y Lana
En el primer caso, Premarck eligió un lenguaje de fichas plásticas de distintos colores y tamaños . Luego del entrenamiento basado en recompensas Sarah aprendió a utilizar estas fichas como palabras. Además, fue capaz de construir correctamente oraciones básicas. Estos ejercicios consistían en completar los "espacios vacíos" de oraciones aprendidas previamente, con palabras semánticamente similares.
El entrenamiento de Lana estuvo basado en el aprendizaje de oraciones básicas mediante un teclado computarizado . El chimpancé seleccionaba la palabra que completaba correctamente la oración presionando las teclas; estas respuestas eran analizadas por el programa de la computadora y en caso de ser aceptadas como correctas la máquina lo proveía de distintos tipo de bebidas o comidas como premios.
En ambos casos se enseñó a los sujetos un stock de oraciones simples y luego se les pedía que completasen espacios en blanco siguiendo reglas sintácticas básicas. Los resultados obtenidos sugerían que los sujetos habían aprendido las reglas que gobiernan las estructuras simples, ya que eran capaces de reemplazar ciertas palabras en las oraciones. Sin embargo, no generalizaban las reglas del modo que se hubiese esperado si la comprensión del significado de las palabras hubiese sido completa.
Proyecto Nim
En los ’70, Terrace llevó a cabo otra serie de investigaciones con chimpancés basadas en la adquisición y uso espontáneo de señas manuales dentro de un grupo social. Mediante la manipulación de sus manos, los monos aprendieron cómo y luego cuándo hacer una seña en asociación con cada uno de una serie de objetos o eventos. El objetivo del trabajo era la producción, es decir el uso de las señas. Mediante técnicas de reforzamiento por premios se le enseñó a Nim cómo indicar una cierta cantidad de objetos. Lo que llamó la atención es que Nim utilizaba cadenas de signos, del tipo de: "dame naranja dame comer naranja". La interpretación de Terrace fue que este encadenamiento de señas no proveía mayor información -en cuanto al nivel de lenguaje adquirido- que el uso de una única seña, y que estas cadenas eran enlazadas entre sí con "cartas múltiples" que siempre resultaban útiles para obtener una recompensa.
Terrance subrayó que las señas gestuales utilizadas eran una imitación completa o parcial de lo que habían visto "decir" a otros en un pasado cercano. De modo que, analizados de esta manera, los resultados de los estudios que inicialmente mostraban la posibilidad de adquisición de lenguaje por chimpancés debían ser reinterpretados. Concluyó que ninguno de éstos había realmente adquirido un lenguaje y, no sólo eso, ninguno sería capaz de adquirirlo.
Se criticó que no se habían hecho tests ciegos que permitiesen estudiar correctamente el aprendizaje de Nim. Dichos tests -en ausencia de entrenadores, para evitar la presión y las expectativas- hubiesen permitido analizar si existía una real comprensión de las palabras utilizadas o si las señas eran sólo el medio para obtener la recompensa.
Por otro lado, científicos que estudiaban la adquisición de lenguaje en niños concluyeron que la imitación puede ser realmente un método normal y efectivo por el cual los niños aprenden y afirman el uso apropiado uso de las palabras. De modo que las imitaciones que realizaban los simios que, según Terrace desacreditaban la adquisición de lenguaje, podrían haber sido parte de la forma en que se adquiere naturalmente.
Proyecto Sherman y Austin
Savage-Rumbaugh utilizó un lenguaje de símbolos en donde cada palabra estaba representada por un dibujo que no tenía ninguna similitud con la imagen del objeto al que hacía referencia. La diferencia fundamental entre este trabajo y los anteriores radicó en el énfasis que se le dio a la comprensión del lenguaje. El uso correcto de los lexigramas seguía siendo recompensado, se modificó la metodología: en lugar de aparear un objeto con su seña y recompensar la respuesta correcta se hizo énfasis en el aspecto comunicativo de los símbolos. Por ejemplo, para enseñarles la palabra "llave" se les daba una caja cerrada. Así, el chimpancé aprendía la palabra relacionándola con su utilidad y no sólo con el objeto material.
Los sujetos aprendieron un gran número de lexigramas, que usaban correctamente para comunicarse con los cuidadores y entre sí. Lo más destacable fue la capacidad que mostraron en la clasificación de palabras. Primero aprendieron, las palabras "comida" y "herramienta" y luego una cantidad de lexigramas que representaban distintas comidas y herramientas; cuando se les pidió que las clasificaran, respondieron perfectamente agrupando las distintas comidas por un lado y las herramientas por otro. Según los investigadores , esto sólo era posible si los chimpancés realmente habían comprendido el significado de cada una de las categorías .
Se cree que eran capaces de categorizar los lexigramas porque el entrenamiento estaba basado en la comprensión; las pruebas requerían el uso coordinado de lexigramas para construir un mensaje o información entre ellos, a diferencia de los demás proyectos en donde, principalmente, se fomentaba la producción de señas o el uso de lexigramas.
Aunque no estaba claro si los demás monos sabían, literalmente, el significado del lexigrama, la ficha plástica o la seña gestual o si simplemente los asociaban con un objeto dado. Como el entrenamiento estuvo basado en la producción del lenguaje, los resultados obtenidos no responden a la pregunta acerca de la comprensión del lenguaje.
Se concluyó que el error radicaba en que estos estudios habían sido construidos al revés; haciendo énfasis en la producción, peroes la comprensión la que funda el lenguaje.
Proyecto Kanzi
En 1990, Savage-Rumbaugh utilizó monos pertenecientes a la especie Pan Paniscus, conocida como bonobo. El primer sujeto estudiado fue una bonobo al que se le trató de enseñar a usar un teclado con lexigramas. Los resultados fueron poco alentadores por lo que se recurrió a un nuevo bonobo con el que continuaría el proyecto cuando tuviese la edad adecuada. El pequeño bonobo fue adoptado por la hembra, acompañándola durante todas las actividades del día, hasta que, a la edad de 2 años fue separado de su madre para comenzar su entrenamiento. Kanzi podía usar signos que había visto anteriormente; no necesitó entrenamiento especial para utilizar el teclado, nombrar objetos ni para manifestar intensiones. Había aprendido mirando a los entrenadores enseñarle a su madre. Se suspendieron los métodos de enseñanza formal y se lo sumergió en un estilo de vida en el que él era parte de la escena social del laboratorio. Se le habló suponiendo que entendía lo que se le decía y, así, el uso de lexigramas acoplado con palabras habladas, sirvió para mostrarle la concordancia entre lo que oía y las símbolos del teclado.
El nuevo enfoque del trabajo asumía que un mono, al igual que un niño, adquiriría el modo dominante de comunicación en el que está inmerso socialmente.
A los 8 años Kanzi poseía una gran comprensión del lenguaje oral. Además, era capaz de diferenciar entre agente, receptor y objeto y de combinar correctamente los lexigramas según reglas gramaticales. Esto último fue puesto a prueba sometiéndolo a distintas clases de requerimientos del tipo: "colocar el objeto X en Y", "dar a S el objeto X", etc., que sólo podían llevarse a cabo si existía un procesamiento de las relaciones sintácticas.
Se demostró que los simios, de manera similar a la de los niños, podrían adquirir lenguaje si se los expone desde el nacimiento a un ambiente donde éste es usado para coordinar actividades sociales. Esto provee de situaciones en la que los sujetos oyen palabras y ven sus respectivos lexigramas usados en contextos variados donde cada uno sirve para predecir eventos en un futuro cercano. Como consecuencia, las palabras adquieren significado y sirven como símbolos para eventos y objetos no necesariamente cercanos.
Se sugiere que, durante la infancia, hay un período sensible para la adquisición de lenguaje . Además, un ambiente lingüísticamente estructurado, en donde el uso del lenguaje posee un rol importante en la vida cotidiana, parecería ser mucho más efectivo para dicha adquisición que el uso de eventos que refuercen la producción.
Esta postura concuerda con la de investigadores del desarrollo infantil que sostienen que el desarrollo del lenguaje es dependiente del desarrollo de la inteligencia, que a la vez depende del crecimiento en un ambiente apropiado; a diferencia de los psicolingüistas, que mantienen que la adquisición del lenguaje se completa rápidamente y es independiente del soporte ambiental y del desarrollo intelectual.
Aunque es imposible argumentar que cualquier animal tiene las mismas capacidades lingüísticas que el hombre, tampoco podría negarse que al menos algunos animales poseen importantes habilidades, entre ellas la comprensión de lenguaje.
Se concluye que las habilidades lingüísticas en simios se desarrollan más eficientemente mediante una crianza temprana en un ambiente lingüísticamente estructurado que mediante en entrenamiento formal a mayor edad. Este hecho implicaría una continuidad entre los simios y el hombre, aún mayor que aquella determinada sólo por la capacidad potencial de adquisición de lenguaje.
EPILOGO
Con relación al momento en que aparece el lenguaje, existen dos posiciones principales. Aquellos que hablan de un origen relativamente reciente, lo correlacionan con la aparición de humanos anatómicamente modernos, con tamaños cerebrales modernos, con áreas de Broca y de Wernicke diferenciadas, y tractos vocales totalmente descendidos . Esto tiene mucho que ver con la idea que la manufactura de herramientas y la producción de lenguaje implican bases cognitivas en común. En este caso, los cambios en la complejidad en la tecnología a través del tiempo también indicarían algo sobre los cambios en las capacidades lingüísticas. La trayectoria de los cambios tecnológicos en la historia homínida presenta dos fases: una muy lenta desde los primeros artefactos, hace unos 2.5 millones de años atrás, hasta unos 200.000 años atrás, donde comienza una segunda fase muy acelerada.
La segunda posición sostiene un origen más temprano del lenguaje, en los comienzos del género Homo. Se sostiene en la expansión cerebral, la configuración humana del cerebro y la aparición de herramientas de piedra en Homo habilis o en los tempranos Homo erectus. Otros investigadores van aún más atrás, situándolo en Australopithecus, en donde comienza el aumento del tamaño cerebral, el bipedalismo y cierta habilidad manual.
El tiempo de aparición del lenguaje tiene para los especialistas implicaciones muy importantes para la naturaleza de la mente. Si apareció recientemente, en los modernos Homo sapiens, entonces el lenguaje fue posterior a otros cambios previos en el cerebro como, posiblemente, un aumento general de la inteligencia, y no hubo mucho tiempo para que el lenguaje influyera en la estructura del cerebro. En cambio, si apareció tempranamente en nuestra evolución, probablemente pasó a través de muchas formas diferentes, y tuvo mayor influencia sobre la evolución de nuestro cerebro. Las múltiples adaptaciones particulares para el lenguaje en el hombre proveen una fuerte evidencia, para algunos, de un origen antiguo para el lenguaje.
Parece haber una relación entre los mecanismos neurales involucrados en el control motor del habla y los responsables de la sintaxis. Asimismo, otras actividades motoras complejas como las responsables de la fabricación de herramientas pueden relacionarse en la evolución de tales mecanismos cerebrales necesarios para el habla y sintaxis. La habilidad para usar palabras con sintaxis sencilla se puede basar en mecanismos neurales.
Se cree que la evolución de las capacidades vocales fue ayudada por gestos manuales. La gesticulación y los cambios en la entonación y en el ritmo aún ayudan a referenciar y organizar las interacciones verbales.
Se acepta que en la evolución de la mente humana actuaron tres elementos interconectados: la expansión del cerebro y de la inteligencia, la conciencia, y el lenguaje.
"El rol del lenguaje en la comunicación evolucionó como un efecto colateral de su rol básico en la construcción de la realidad" Jerison. Es decir, según este autor, el lenguaje es un medio de construir imágenes mentales.
MÉTODOS UTILIZADOS EN LA RECONSTRUCCIÓN DE LA VIDA HOMÍNIDA
Evidencias fósiles
Hay 3 tipos principales: los fósiles en sí mismos, los indicios arqueológicos, y el contexto ambiental en el cual esos homínidos vivieron.
Fósiles.
Los huesos fosilizados proveen información sobre el esqueleto y sobre sus modificaciones durante el crecimiento, las tensiones biomecánicas, y los cambios causados por la dieta y las patologías.
A partir de los fragmentos de los cráneos homínidos puede estimarse el volumen total del cerebro. El tamaño se incrementó a lo largo del tiempo.
Mediante el uso de endocastos (impresiones hechas por los pliegues de la corteza cerebral sobre el interior del cráneo) se han inferido cambios en la organización del cerebro, tales como la expansión de áreas sensoriales y de áreas involucradas con el control motor o con el lenguaje. Estimaciones de las relaciones en el tamaño corporal permiten hacer inferencias sobre parámetros ecológicos.
Estudios hechos a partir de huesos fósiles sobre el sexo, la edad, la tasa de desarrollo y las patologías también iluminan aspectos importantes del comportamiento. En primates, como en muchos mamíferos, un mayor tamaño corporal de los machos, comparados con las hembras, refleja la competencia entre machos observada en sistemas sociales poligínicos (un macho acaparando varias hembras), en los que existe más variación en el éxito de apareamiento para los machos que en sistemas monogámicos. El dimorfismo sexual en el tamaño del esqueleto entre los homínidos también nos da pistas sobre su vida social. Tal dimorfismo es relativamente grande en los primeros homínidos y se reduce en los Neanderthalenses, aproximándose al nivel que se observa en los humanos modernos.
Los humanos nacen inmaduros y continúan su maduración durante un período, esto se relaciona con unos patrones complejos de comportamiento, tales como un aumento en la dependencia del aprendizaje y de los cuidados parentales, y en la defensa de un hogar base.
Restos arqueológicos
Los registros arqueológicos comienzan cuando los homínidos comenzaron a manufacturar artefactos con materiales duros como piedras trabajadas, artefactos, huesos de animales o restos de plantas.
Las herramientas de piedra proveen información a pequeña escala sobre patrones de movimiento y uso de la tierra, y también información sobre el comportamiento, en la medida en que los métodos de manufactura pueden indicar las capacidades de las manos y del cerebro.
Contexto geológico
La información geológica ayuda a la interpretación de los restos arqueológicos, y también puede iluminar el contexto ecológico en el que vivieron los primeros homínidos. El estudio de los sedimentos permite hacer inferencias sobre la distribución de caracteres físicos, tales como lagos, costas y planicies, y los análisis geoquímicos, junto con los fósiles de plantas y animales, dan información sobre las fluctuaciones climáticas, la vegetación, y las comunidades ecológicas en las que vivieron los homínidos.
Las extrapolaciones más importantes provienen de la observación de dos líneas principales: civilizaciones contemporáneas y animales cercanamente emparentados con nosotros.
Civilizaciones contemporáneas
Ciertos estudios de evolución humana utilizan como analogía la búsqueda de paralelismos entre el registro arqueológico y el etnográfico (el estudio de la forma de vida de civilizaciones contemporáneas), a partir de los cuales se pueden hacer inferencias sobre aspectos de la prehistoria. El supuesto que subyace a este método es que el material arqueológico observable de una cultura está correlacionado con otros aspectos de la organización social y económica. Sin embargo, esta técnica presenta algunos inconvenientes. Para empezar, las fuentes de información etnográficas son muy diversas y no igualmente confiables. Incluso teniendo en cuenta este hecho, el registro etnográfico puede estar sesgado, al igual que el arqueológico, por la preservación. El registro etnográfico es una muestra del comportamiento humano, no una verdadera representación de su patrón general de variación.
Otro problema tiene que ver con la diversidad que presenta el comportamiento moderno.
Analogías animales
Los problemas que enfrenta el paralelismo etnográfico, en cuanto a la variedad y diversidad del comportamiento y la cultura humanos, se relacionan con la controversia que existe sobre si es posible establecer modelos generales y predictivos de comportamiento humano.
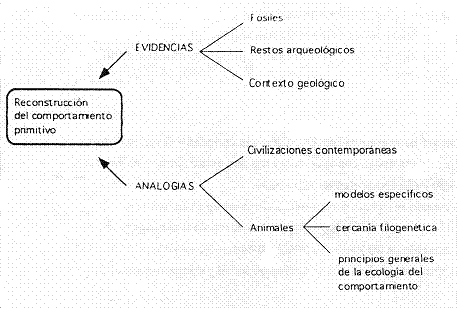 Puede esperarse que homínidos de un millón de años atrás o más hayan tenido
patrones de comportamiento que se parezcan más a los de primates no humanos
que a los de humanos modernos. El comportamiento, la organización social y
la ecología de los primates no humanos deberían ser una mejor guía para
entender las adaptaciones de los primeros homínidos, mientras que su
anatomía puede reflejar similitudes funcionales.
Puede esperarse que homínidos de un millón de años atrás o más hayan tenido
patrones de comportamiento que se parezcan más a los de primates no humanos
que a los de humanos modernos. El comportamiento, la organización social y
la ecología de los primates no humanos deberían ser una mejor guía para
entender las adaptaciones de los primeros homínidos, mientras que su
anatomía puede reflejar similitudes funcionales.
Los primates modernos han sido usados para modelar el comportamiento de nuestros antecesores de acuerdo a tres vías principales:
1. Identificar una especie en particular que aparezca como similar en algunas características básicas de los homínidos, y buscar en ella patrones de comportamiento que puedan transferirse a aquellos.
2. Guiarse por la filogenia y buscar sólo en los simios africanos capacidades o comportamientos compartidos que puedan haber estado presentes en un antecesor común del hombre y los simios.
3. A partir del estudio de la ecología del comportamiento, inferir principios generales que puedan aplicarse a los homínidos.
Elección de las analogías correctas
Los modelos de analogías son útiles, ya que pueden ayudar a encontrar las similitudes y las diferencias entre el presente y el pasado. No obstante, como el pasado sólo es accesible a través de fósiles, es importante que los modelos sean comparados directamente con el registro fósil.
La tafonomía estudia cómo el comportamiento de forma vivientes se puede manifestar como fósiles.
Semana 11
El origen y evolución del lenguaje. Teórico.
Hay dos modelos en cuanto a la origen y evolución del lenguaje, por un lado, los conductistas con Skinner, los cuales argumentan que aprendemos a hablar de la misma forma que aprendemos cualquier otra habilidad. Los niños son recompensados cuando hablan correctamente y son reprobados cuando cometen errores. Nosotros podemos hablar, mientras los chimpancés no pueden, porque somos mejores aprendiendo, no hay nada especial acerca del lenguaje.
Contrariamente, encontramos a Chomsky y seguidores sostienen que los humanos tienen una competencia peculiar para el lenguaje que no es solo un aspecto de su inteligencia general. Aprendemos a articular, y a comprender un número indefinidamente extenso de oraciones gramaticales y evitar un número aún mayor de oraciones no gramaticales. Por lo tanto, difícilmente podamos aprender que oraciones son gramaticales y cuales no por ensayo y error. Por el contrario, debemos aprender las reglas que generan las oraciones gramaticales. Estas reglas son de una gran sutileza, entonces, aunque las adquirimos y aplicamos no las podemos formular explícitamente.
Es difícil creer que podemos dominar tan fácilmente tales reglas a menos que estemos genéticamente predispuestos para hacerlo. De igual forma, otra razón para pensar que no podemos aprender a hablar por ensayo y error reside en la pobreza del input (entrada de información lingüística) en el que un niño debe confiar. Luego de escuchar un juego definido de articulaciones el niño aprende a generar un número mayor indefinido de oraciones gramaticales. Esto implica que el niño aprende reglas y no solamente un juego de oraciones. La habilidad para aprender a hablar es independiente de la inteligencia general.
Es difícil identificar qué *es lo innato. No es la habilidad de hablar un lenguaje particular, pero si la capacidad de hablar cualquier lenguaje. En comunicación, el significado es convertido en una secuencia lineal de sonidos (o gestos en un lenguaje de sordomudos). Chomsky se refirió a esta secuencia como la “estructura de la superficie”. La linealidad no es necesariamente un rasgo de los sistemas de comunicación: nuestros mensajes pueden ser multicanales o puede consistir en despliegues en dos direcciones. Pero, antes de comunicarse hay que tener algo que decir, hay que pensar. Hay que ser capaces de representar el mundo en nuestras mentes y manipular la imagen que hemos formado: el pensamiento requiere gramática mental.
La evolución de la competencia lingüística requirió la adquisición de dos habilidades. La primera, requerida tanto para el lenguaje como para el pensamiento, es la habilidad de formar representaciones complejas del mundo en nuestras mentes. La segunda, más específica de la comunicación es la habilidad de transformar estas representaciones en una secuencia lineal de símbolos.
Hay dos analogías entre el rol del lenguaje en el origen de la sociedad humana y las transiciones tempranas de la evolución. La primer analogía fue señalada por Jablonka donde el origen de los organismos multicelulares requirió un segundo sistema de herencia, basado en la metilación y en otros medios de etiquetado de ADN. Esto provocó una división del trabajo entre las células y puede haber ayudado a juntar grupos de células, de esta forma el organismo se convirtió en el blanco de la selección, y no la célula aislada. En la sociedad humana, el lenguaje constituye el segundo sistema de herencia. Esto posibilita la división del trabajo, y une a los grupos humanos a través de los mitos y el ritual.
La segunda analogía corresponde a Chomsky el cual expresó que la característica esencial del lenguaje es que se pueden articular un número indefinido de oraciones con significado. El origen de la vida requirió replicadores que pudieran existir y ser copiados en un número indefinido de formas.
LENGUAJE Y REPRESENTACIÓN
El mono vervet de África del Este emite tres llamadas de alarma: una para las serpientes pitón, otra para las águilas marciales y otra para los leopardos. Cuando un mono ve a alguno de estos animales emite una llamada y provoca la respuesta adecuada en los monos que la escuchan: por ejemplo, la llamada para el leopardo hace que los monos se trepen en lo alto de un árbol, respuesta que sería inadecuada si el predador fuera un águila. Se sugirió que el lenguaje se puede rastrear (buscar sus orígenes) hasta un sistema de alarmas de este tipo. Esto implica que la señal de alarma es en efecto una palabra tal como serpiente pitón, la llamada es un signo de la serpiente.
Los estudios de habituación en cuanto a la llamada de los monos descubrieron que si una señal de alarma no es acompañada por una amenaza objetiva los animales se acostumbran y dejan de reaccionar. Las llamadas wrr y chatter (diferenciadas acústicamente), son usadas por los monos para llamar la atención sobre la presencia de otros grupos de monos. Demostraron que generalizan la habituación teniendo en cuenta el significado de la llamada, de forma que un sujeto habituado al Wrr de un sujeto X, cuando escucha un Chutter del mismo sujeto sigue dando una respuesta habituada como si estuviera escuchando un Wrr. Sin embargo, estos monos identifican individuos. Se comprobó que un mono que se habituó al sonido wrr de otro mono generaliza la habituación al chatter del mismo individuo, pero no al chatter de los otros. Seyfarth y Cheney concluyeron que estas llamadas son representaciones semánticas. El uso del lenguaje en los vervets es sin embargo bastante limitado. En particular, un vervet no usa una llamada para referirse a un objeto que no esté presente aquí y ahora.
Russell argumentó que hay un buen correlato entre los objetos “allá afuera” y las palabras, o señales de alarma en estos monos. El problema es que la correlación no es directa. Primero hay una correlación desde el objeto serpiente al concepto “serpiente” y luego desde “serpiente” a la palabra serpiente, tal como de Saussure lo señalaba. Los vervet deben tener un concepto de serpiente en sus cerebros, de otra forma no las podrían reconocer. De hecho, los monos jóvenes tienen que aprender el rango semántico de sus llamadas.
Para los humanos un sistema de representación potencialmente completo es decisivo. Surge la analogía sobre que el origen de la vida fue marcado por el pasaje de replicadores hereditarios limitados a ilimitados. Ahora parece que el origen de la vida humana fue marcado por el pasaje de representaciones semánticas limitadas a ilimitadas.
Los humanos usan el lenguaje para la comunicación, pero bien puede ser que el aspecto más importante del lenguaje sea usado para la representación interna en el cerebro .
Pensamiento, Lenguaje y habilidades asociadas.
Se puede distinguir una serie de habilidades asociadas al lenguaje necesarias para el pensamiento como la formación de conceptos (categorización y jerarquización), la predicación y el establecimiento de relaciones entre conceptos.
1. Conceptos: Categorización y Jerarquías.
Los animales pre-lingüísticos pueden tener un sistema de representación. Las representaciones llevan a la formación de categorías. La formación de categorías es la agrupación de todas aquellas representaciones que requieren de una misma respuestas comportamental. Esto es independiente de, si las representaciones son o no innatas (independiente de la experiencia). Una categoría implica necesariamente el manejo de un concepto: esto significa que a los animales se les debe otorgar la facultad de formación de conceptos. El lenguaje solamente proporcionó etiquetas para conceptos que se derivan de una experiencia pre-lingüística.
Las categorías se pueden agrupar de manera jerárquica, particularmente si son asociadas con palabras. Todos los ítems léxicos son parte de una estructura jerárquica.
2. Predicación:
Una característica fundamental del lenguaje es la distinción entre sujeto y predicado. Ya que muchas cualidades se pueden predicar de cada entidad, el rango de cosas que se pueden decir con un vocabulario limitado es mayor de lo que sería si tuviéramos una palabra separada para cada párrafo, entidad + cualidad. La distinción sujeto-predicado es una característica universal del lenguaje y es la base de nuestra capacidad de decir y comprender un número indefinidamente largo de oraciones .
3. Elementos gramaticales y la relación entre conceptos:
El pensamiento humano requiere la capacidad de la formación de categorías ordenadas jerárquicamente y la predicación junto a la formación de representaciones estructuradas del mundo que puedan ser manipuladas en nuestras mentes. Debemos poder ordenar los conceptos en un patrón y luego alterarlo. Esta última habilidad es análoga a la gramática, como lo demuestra el uso de las palabras entonces, si entonces, dentro y debajo, al describir el pensamiento expresado anteriormente.
Las palabras puramente gramáticas como “por lo tanto” y “si” que no tienen referente demostrable representan más o menos la mitad de las palabras en el lenguaje humano. También se incluyen las inflexiones (-ando, -endo, -ado, -ido). Los sustantivos pueden ser singulares o plurales, la mayoría de los verbos indican el tiempo presente o pasado, y así sucesivamente. Los ítems gramaticales mantienen unidas las partes más importantes de la oración. Indican el número relativo, el tiempo relativo, la dirección relativa, la familiaridad relativa, la posibilidad relativa, la posición, la necesidad, la existencia, etc. Aunque, dichos sistemas gramaticales sólo se refieren a un número pequeño de relaciones, aunque muchos lenguajes pueden proporcionar distinciones que no son posibles en otros idiomas.
Los ítems puramente gramaticales son necesarios para el lenguaje y para el pensamiento, el proceso de formación de conceptos, predicación y el reconocimiento de relaciones entre los conceptos. Las habilidades mentales necesarias que pueden haber evolucionado en la primera instancia para el pensamiento más que para la comunicación o al menos para pensar tanto como para comunicarse.
La comunicación requiere una representación adecuada del mundo en el cerebro y que esta representación se convierta en una secuencia lineal de sonidos o, viceversa, que una secuencia lineal de sonidos se convierta en una representación en el cerebro.
CARACTERÍSTICAS DE LA SINTAXIS
Oxford define a la sintaxis como el orden de palabras en el cual estas trasmiten el significado colectivamente mediante su conexión y relación.
La moderna visión de la sintaxis y de la característica innata de la gramática universal es debida a Chomsky quien fue el primero en distinguir semánticamente entre oraciones sin sentido gramatical tales como Ideas verdes descoloridas duermen furiosamente (en oposición a Ideas nuevas revolucionarias ocurren infrecuentemente), y en oraciones incorrectas desde el punto de vista sintáctico tales como Nuevas ocurren revolucionarias infrecuentes ideas.
a)
 Estructura Universal de la Frase (Bickerton)
Estructura Universal de la Frase (Bickerton)
X representa cualquier categoría léxica (sustantivo, verbo, adjetivo, preposición, etc) que puede expandirse para formar una frase. La n luego de X´ indica que este nivel, a diferencia de X y X´se puede repetir. Los paréntesis alrededor del especificador y del complemento indican que estos items no son obligatorios. las flechas bidireccionales indican que las posiciones relativas de los items se pueden intercambiar.
b) Estructura de Argumento.
El conocimiento de la estructura de los argumentos es innato. Lo innato se refiere a la prescindibilidad de la experiencia, pero, al mismo tiempo, podemos proponer que disponemos de una particular habilidad por la cual, a pesar de no ser instruidos formalmente a cerca de la regla, podemos categorizar los verbos una vez conocido su concepto y no equivocarnos en el número y tipo de argumento que le corresponde.
c) Elementos Nulos:
Una tercera característica es la habilidad para alterar el significado de las oraciones al mover las palabras. Cuando mueven las palabras mantenemos el rastro de ellas, hay un “elemento nulo” presente en la oración. Por ej.: ¿Qué has visto? El elemento nulo sería el sujeto al quien se le hace la pregunta.
ADQUISICIÓN DEL LENGUAJE
La razón más importante para pensar que los humanos tiene una competencia innata para aprender lenguajes es que los niños adquieren un dominio de reglas gramaticales complejas al escuchar a otros hablar.
Los niños logran desarrollar un lenguaje con un buen manejo del intrincado sistema de reglas, antes de comenzar sus estudios primarios, en los cuales van a recibir sus primeras instrucciones en esta y otras habilidades cognitivas. Todos los niños de una comunidad logran adquirir una gramática semejante, antes de comenzar sus estudios primario, independientemente de la gran variabilidad en la estimulación experimentada. La universalidad del lenguaje es tal que comparada con otras tareas cognitivas (cálculos aritméticos) su adquisición resulta sorprendentemente independiente de la calidad y cantidad del Input.
SELECCIÓN NATURAL PARA EL LENGUAJE
Chomsky ponía un gran énfasis en la característica innata de los dispositivos de adquisición del lenguaje. Dijo que, aunque no tengamos un “órgano del lenguaje”, especular acerca de sus orígenes es tan inútil como hacer lo mismo acerca de cualquier otro órgano.
En esta sección discutimos si el origen del lenguaje se puede explicar por la selección natural siguiendo el tratamiento de Pinker y Bloom.
Partimos de la presunción de que la selección natural es la única explicación posible para un diseño adaptativo. Siguiendo aGould y Lewontin, se podría suponer que es un spandrel: un subproducto no seleccionado, resultado de un diseño propicio para algún otro propósito. (Un spandrel es específicamente una cavidad (no intencional), producto de una estructura particular de arcos en arquitectura). Más específicamente, el lenguaje podría ser un spandrel modificado o no modificado.
Si el lenguaje está modificado, la selección natural sería la fuerza modificadora. La única alternativa interesante no seleccionista sería que el lenguaje es un spandrel no modificado, una preadaptación perfecta. No se ha construido una teoría seria a lo largo de esta propuesta.
Hay habilidades mentales que son productos no seleccionados de la capacidad cognitiva. Si se piensa que la competencia lingüística fue sólo una consecuencia de un aumento general de la inteligencia, entonces podemos tratarla como un spandrel, es decir un trazo cuyo origen no requiere explicación. Esto es lo que Chomsky niega, insisten en que el lenguaje es un caso particular. Entonces, la competencia lingüística tal como la habilidad para formar y manipular representaciones mentales evolucionó gracias a la selección de la habilidad cognitiva general; y la competencia específica para convertir el significado en estructura superficial evolucionó gracias a la selección de la habilidad comunicacional.
Piatelli-Palmarini plantearon que, debido a que la gramática tiene varios elementos arbitrarios esta no constituye una adaptación predecible para la comunicación, entonces carece de diseño y no puede ser un producto de la selección natural. Esta opinión no comprende la naturaleza de la evolución por selección natural: en general los resultados no son predecibles.
Lieberman planteó que la gramática universal no puede ser producto de la selección natural porque no hay variación genética para ella. Pero hay una variación genética ya que no hay una justificación clara para asegurar que no hay variación genética.
Dunbar evidenció que para la selección de las habilidades sociales podría haber sido el motivo principal para el incremento del tamaño del cerebro de los primates. En los humanos hubiera sido selectivamente ventajoso comunicar acerca del tiempo, de la posesión, de las creencias, de los deseos, de las tendencias, las obligación, la verdad, la probabilidad, las hipótesis y los contraargumentos. La carrera armamentista intelectual tuvo lugar dentro de las especies mismas.
Dificultades :
1) Una “mutación” para una mejor sintaxis no se generalizaría porque al principio no podría ser comprendida . Una respuesta parcial a esta dificultad puede darse al señalar la generalización de nuevas palabras.
La respuesta a la dificultad aparece en el proceso de “asimilación genética”. Como lo señaló Waddington en 1956 una característica que primero aparece como respuesta a un estímulo ambiental puede asimilarse genéticamente si es ventajosa desde el punto de vista de la selección y aparecer sin el estímulo. Este autor propuso el efecto llamado asimilación genética.
El proceso funciona porque las elección favorece a los genotipos que responden más rápidamente a los estímulos. Hinton y Nowlan simularon la asimilación genética. Imaginaron una tarea que inicialmente debe ser aprendida mediante ensayo y error pero que gradualmente se convierte en una tarea programada genéticamente y realizada sin aprendizaje. La población no puede evolucionar en la habilidad para realizar la tarea solo mediante selección natural actuando sobre mutaciones azarosas sin aprendizaje individual, porque muchas mutaciones se deben adquirir simultáneamente, pero la evolución es exitosa si hay una combinación de aprendizaje y selección natural.
Pinker y Bloom plantearon que la evolución del lenguaje es el campo ideal para la operación de tal proceso. Nosotros aprendemos nuevas habilidades lingüísticas, pero también sabemos que muchas habilidades lingüísticas se asimilan genéticamente.
2) Otra dificultad es que la gramática debe operar como un todo: los cambios pueden tener consecuencias drásticas. Esto implica un origen saltasionista de la gramática según escribió Bates. Si los principios estructurales básicos del lenguaje no se pueden aprender (de abajo hacia arriba) o trasmitir (de arriba hacia abajo), sólo hay dos explicaciones posibles para su existencia. O bien la gramática universal nos fue confiada directamente por el Creador, o bien nuestra especie sufrió una mutación de magnitud sin precedentes, un equivalente cognitivo del “big bang”.
La mayoría de las objeciones expresadas por los lingüistas a los posibles intermediarios se limitan a decir “si la característica X de la gramática no existiera no se podría decir Y”. Esto es generalmente verdadero e irrelevante. El problema no es si se puede decir Y, sino si la gramática sin X sería mejor que la ausencia de gramática.
USO DE HERRAMIENTAS Y LENGUAJE: COMPORTAMIENTO SECUENCIAL ORGANIZADO JERÁRQUICAMENTE.
Greenfield en 1991 sugirió la relación entre usos de herramientas y desarrollo del lenguaje. El primer punto para establecer es que hay una similitud formal entre “la gramática en acción” y el protolenguaje. El desarrollo de la gramática en acción se puede ilustrar mediante la forma en que los niños manipulan un juego de tazas (poner una dentro de otra de mayor a menor). Los niños pasan a través de tres etapas de las cuales sólo en la tercera, la estrategia de ensamble es jerárquica. En esta estrategia, una estructura (una taza dentro de la otra) que fue producida por una acción se trata como una unidad en una segunda acción.
También es el caso que, tanto en la gramática de acción como en la construcción de oraciones los niños tienen las estrategias sucesivamente en el mismo orden. Sin embargo, es posible que se lleven a cabo mediante diferentes estructuras neurales ya que no se desarrollan simultáneamente.
Greenfield sin embargo sugirió que el verdadero paralelismo es el que existe entre la gramática de acción y la construcción de palabras.
Los niños pasan a través de las siguientes etapas en la construcción de palabras:
i. Las primeras palabras, hacia el final del primer año, son sílabas de vocales y consonantes reduplicadas, tales como dadá y mamá, en las cuales una única consonante se combina repetidamente con la misma vocal. A esta edad los niños tocan repetidamente un objeto con otro.
ii. Una única consonante se combina con una única vocal, como es na (para no). A esta edad un objeto se une a otro objeto; como la estrategia de formación de pares para las tazas apiladas.
iii. Una única consonante se combina sucesivamente con dos vocales diferentes, como en teta, en esta etapa los niños combinan la misma taza con otras dos tazas diferentes, pero sin combinar tres tazas juntas.
iv. La consonante inicial varía, pero la vocal permanece constante como en casa, masa (Papá “casa”). Esto corresponde a la estrategia de “recipientes” para las tazas apiladas.
v. Finalmente se combina el desarrollo silábico ensamblado como en tren.
La evidencia para una conexión entre dos procesos se deriva de las afasias (defectos del lenguaje) que resultan de daños en el área de Broca, localizada en el lóbulo frontal izquierdo de la corteza cerebral. Los adultos que sufrieron daños en éste área son incapaces de producir discursos organizados sintácticamente: la estructuración de su lenguaje carece de rastros de estructura de árbol. A menudo muestran una falla similar en la manipulación de objetos. El área de Broca no sólo está comprometida en la sintaxis sino también en la fonología. Aunque la incapacidad para manejar jerarquías se asocia con lesiones en el área de Broca y no de otro tipo de lesiones, hay complicaciones. La asociación entre objetos y manipulación de oraciones es más fuerte en los niños que en los adultos: algunos adultos son buenos para la sintaxis y malos para la gramática de acción y viceversa. Greenfield sugiere que al comienzo del desarrollo ambas funciones son realizadas por el área de Broca, pero posteriormente las funciones se separan y son asumidas por un área de la región prefrontal.
Una idea del máximo dominio de la sintaxis logrado por animales entrenados lingüísticamente en cautiverio, es proporcionada por el estudio de Herman de dos delfines nariz de botella, Phoenix que fue entrenada para responder a señales acústicas y el otro Areakamai para gestos. Sólo se desempeñaron como receptores de señales, no como emisores. Phoenix tenía un vocabulario de treinta (30) palabras. Ella aprendió a distinguir entre verbos con uno o dos argumentos. Ambos delfines respondieron a oraciones de 2 a 5 palabras. Los resultados:
· Utilizan la estrategia de ensamble; por ejemplo, tratando caño de la superficie como si fuera una unidad.
· Utilizan el orden de las palabras para indicar significados; por ejemplo, pelota traer caño y caño traer pelota son interpretadas diferentemente. Se les enseñó a interpretar el orden de las palabras en forma diferente. Por ejemplo, Areakamai respondía a la instrucción caño pelota traer (y no caño traer pelota) para traer la pelota al caño. Descubren el significado de una oración analizando la estructura completa, y no simplemente procesando de derecha a izquierda, entonces las palabras posteriores pueden modificar el significado de las palabras anteriores: por ejemplo, para Areakami la oración cesto sobre, la palabra cesto es objeto directo mientras que en cesto pelota traer, cesto es una meta.
· Los delfines eran capaces de comprender las oraciones nuevas y las formas sintácticas.
· Hay una evidencia preliminar de que los delfines pueden comprender oraciones conjuntas.
No se puede afirmar que los animales no tienen competencias sintácticas. Sin embargo, su capacidad se demostró al recibir señales, no al emitirlas y esto fue el logro de animales cautivos con entrenamiento lingüístico. Tenemos poca idea acerca del funcionamiento de la comunicación en estado de libertad.
DAÑO CEREBRAL Y DESÓRDENES DEL LENGUAJE
En cuanto al daño en el área de Wernicke, los pacientes articulan un lenguaje sin sentido, pero sintácticamente correcto. Su manipulación de los objetos sufre de la misma incapacidad para recordar el contenido a pesar de la estructura correcta.
La capacidad para nombrar los conceptos requiere tres habilidades : formar los conceptos, formar nuevas palabras y formar conexiones entre palabras y conceptos.
Los pacientes con daños en elsegmento temporal del gyrus lingual izquierdo sufren de “ anomia del color”. Experimentan el color normalmente y están capacitados para la morfología de las palabras, pero son incapaces de unir los nombres a los colores. El lazo entre la palabra y el concepto está dañado.
Los pacientes con daño en lascorteza medio temporal y anterior reconocen el color, pero no pueden nombrarlos. Esta inhabilidad para nombrar cosas es más pronunciada para los artefactos naturales que para los humanos, sugiriendo que el cerebro puede usar diferentes partes para representar objetos pertenecientes a estas categorías.
Pacientes con daño en lóbulo temporal anterior izquierdo pueden perder la habilidad para recordar los nombres de personas.
Cada evidencia para una asociación entre habilidades lingüísticas particulares y regiones particulares del cerebro reposa en la visión de que la competencia lingüística es específica y no simplemente un aspecto de la habilidad cognitiva general.
LA GENÉTICA DE LOS DESORDENES DEL LENGUAJE
Gopnik describió a una familia de habla inglesa en la cual ocurre un desorden particular del lenguaje (disfasia), en 16 de 30 miembros de la familia, a lo largo de tres generaciones. Los datos son consistentes con la visión de que los desórdenes son causados por un gen autosomal dominante. La interpretación genética se refuerza por el hecho de que algunos miembros de la familia pueden estar afectados mientras que otros son normales: esto permite descartar la idea de que el rasgo es familiar debido a que los niños lo aprenden de sus padres. Los difásicos permanecen con este desorden aun cuando reciben entrenamiento lingüístico especial y aun cuando un padre es normal. No hay indicación de que otras habilidades cognitivas estén afectadas: estas personas pueden hacer bromas y conversar y pueden ser muy aptos para las matemáticas. No hay fallas generales para manejar estructuras jerárquicas.
Los niños pueden aprender ejemplos particulares de singulares y plurales, así como de tiempos de verbos, tal cual todos debemos aprender los significados de palabras particulares como “caballo” o “vaca”, pero no pueden generalizar.
Este caso tiene algunas implicancias importantes. En primer lugar, aunque las personas afectadas de esta familia están disminuidas, no carecen de gramática. Están bastante mejor de lo que estarían si no pudieran hablar en absoluto. En otras palabras, puede haber intermediación entre una competencia perfecta y la carencia de esta. En segundo lugar, el impedimento es específico del lenguaje: no hay defecto mental. Esto confirma la visión de Chomsky de que la competencia lingüística no es un mero derivado de la inteligencia general .
PROTOLENGUAJE
Hay cuatro fuentes que pueden indicar lo que fue el lenguaje primitivo: 1) El lenguaje de los simios, 2) El lenguaje temprano de los niños, 3) El lenguaje del niño “Casper Hauser” (aislado de toda estimulación con un lenguaje) y 4) Las jergas.
1) El lenguaje de los Simios y 2) El lenguaje temprano de los niños.
Tanto los niños en la etapa de dos palabras como el chimpancé entrenado Washoe (Gardner y Gardner) formulan sentencias que ilustran la atribución de cualidades a objetos, la posesión de objetos no vivientes por seres vivientes, la localización de las acciones, la localización de agentes y pacientes en las acciones. Los dos sets son formalmente idénticos. No hay elementos gramaticales ni hay rastros de estructura. La única diferencia real entre ambos es que los niños parecen categorizar por el hecho de categorizar, mientras que los simios toman sólo objetos que quieren o de acciones que quieren realizar o han realizado: los niños tienen más curiosidad acerca del mundo que los rodea.
3) El lenguaje del niño “Casper Hauser”
“Genie” fue una niña de 13 años encontrada en California en 1970. Su padre la mantuvo aislada en su cuarto, sola, cuando tenía un año y medio. Era incapaz de hablar. Aunque era normal su capacidad para formar conceptos, permaneció detenida en el nivel lingüístico. Genie adquirió lo que se llama protolenguaje. Teniendo en cuenta la similitud con el lenguaje de los simios, el protolenguaje sería filogenéticamente antiguo. No hay aparentemente períodos críticos en los cuales se deba adquirir.
4) Las jergas
Las jergas se desarrollan cuando la gente debe comunicarse con otra gente sin compartir el lenguaje común, y sin un lenguaje que sea un medio obvio. A veces la jerga permanece como un nivel primitivo a través de varias generaciones. Lo notable acerca de la jerga es la escases de conocimientos lingüísticos que sus hablantes tienen que emplear para comunicarse a través de las barreras del lenguaje
La diferencia crucial entre varias formas de protolenguaje y el lenguaje verdadero son:
- El orden de las palabras en el protolenguaje no se rige por reglas sintácticas formales .
· En el lenguaje ordinario, los elementos nulos indican los sitios de la oración, donde podemos inferir que algún componente esta imaginariamente presente. En el protolenguaje, cualquier elemento puede faltar de cualquier posición de una forma impredecible.
· La estructura de la frase está casi ausente. Las pocas excepciones aparentes pueden haber sido aprendidas, como ítems léxicos y frases idiomáticas.
- Hay pocos ítems gramaticales.
DEL PROTOLENGUAJE AL LENGUAJE
Los niños al aprender la jerga de los adultos crean una jerga transformada descrita como la transición de la jerga al criollo (Creol). Los lenguajes Criollos surgen cuando los niños de los inmigrantes no fueron expuestos a ningún lenguaje natural, sino sólo a la jerga.
Mientras que casi todos hablan su propia jerga, las reglas del Criollo son uniformes de hablante a hablante. Uno debiera esperar que la gramática provenga inminentemente de un lenguaje fuente o que haya una mezcla derivada de todos los lenguajes fuentes disponibles, pero no es así.
Una característica del Criollo es que permiten el doble negativo “No quiero nada de zapallo”. Una segunda característica común es distinguir oraciones de preguntas sólo por la entonación. Una tercera característica gramatical compartido por varios lenguajes Criollos es el método de conjugar verbos. Bickerton llamó la atención que, entre las edades de 2 a 4 años, los niños que aprenden el inglés a veces cometen errores que podrían ser correctos gramaticalmente en Criollo: por ejemplo, los niños a veces prueban con el doble negativo.
CONLCLUSIONES
Los humanos se diferencian de los simios no solo en su competencia con la gramática, sino también en su habilidad para producir y percibir sonidos. Una de las dificultades en enseñar a hablar a simios, reside en su incapacidad para producir gran parte de los sonidos usados en el habla humana. La evolución de los humanos requirió, por un lado, de cambios anatómicos y por otro de un mejoramiento en los mecanismos cerebrales responsables de la producción y percepción de sonidos. Anatómicamente, el descenso de la laringe en humanos incrementó el rango de posibles sonidos a producir, así como el costo de un significativo aumento en las probabilidades de atragantarse cuando comemos o tomamos.
Somos capaces de producir y percibir segmentos de sonidos a una velocidad de 25 por segundo. Esta rápida transmisión es necesaria: sin ella, nosotros olvidaríamos el comienzo de una sentencia antes de que esta hubiera terminado.
Desarrollo Individual: Análisis de la ontogenia del comportamiento. (Teórico)
Durante el desarrollo de un individuo, desde la formación del huevo fertilizado -hasta su muerte, los seres vivos pasan por distintos estadios, cada uno caracterizado por diferentes rasgos morfológicos, fisiológicos, comportamentales.
Este proceso de cambio denominado es Ontogenia.
En sus orígenes la principal disputa teórica acerca de la naturaleza del proceso ontogénico involucró a dos corrientes de pensamiento bien diferenciadas "Los Preformistas y los Epigenetistas".
Los Preformistas consideraban que una versión en miniatura del individuo adulto ya estaba presente en el cigoto (óvulo fecundado) y que ninguna parte del animal se crea antes que otra sino simultáneamente.
Por el contrario, los epigenetistas proponían la existencia de transformaciones durante el desarrollo que hacían pasar a los organismos en desarrollo por distintos estadios caracterizados no solo por crecimiento sino también por diferenciación. Los trabajos de embriología del pollo realizados por Wolff (1733-1794) permitieron confirmar las ideas Epigenetistas, las cuales, a grandes rasgos, son aceptadas hoy en día.
Dos nuevos marcos teóricos de discusión hicieron su aparición en distintos momentos, uno producto de interpretaciones diferentes de las ideas epigenetistas denominada controversia Natura-Nurtura o dicotomía innato-aprendido. La otra disputa está basada en la relación entre la filogenia (historia evolutiva de las especies) y la ontogenia (historia evolutiva del individuo).
El Proceso Ontogénico y la controversia Natura-Nurtura:
Mendel propuso que los factores internos (genes) eran los causantes del aspecto o forma de los organismos con vida. Darwin consideraba que los factores externos (el ambiente) eran los responsables de dar forma a los organismos y los causantes de la diversidad biológica. Así, durante un período, el desarrollo fue concebido como la expresión de rasgos latentes en los genes, los cuales necesitaban de un ambiente adecuado para poder actuar. El ambiente era el encargado de condicionar el normal desarrollo del individuo, y solo ciertos individuos estarían capacitados para desarrollarse adecuadamente en un medio particular (Selección Natural).
Esta visión de genes determinando los rasgos del individuo y ambiente responsable de determinar a las especies ha sido modificada por una hipótesis donde la interacción entre los genes, los organismos y el ambiente es reciproca .
Los machos y hembras (animales no humanos) poseen diferencias genéticas marcadas en sus cromosomas sexuales siendo los machos XY y las hembras XX. El cromosoma "Y" posee genes diferentes a los del "X" entre los cuales existirían uno o más, cuyo producto sería responsable al menos del comienzo de la masculinización del sujeto en desarrollo. Dicho producto es denominado "Factor Determinante de Testículos", el cual organiza el desarrollo de los testículos en el embrión macho. Las células testiculares que comienzan a proliferar en número secretan testosterona (Hormona esteroide masculina), la cual posee un efecto organizativo a dos niveles. Por un lado, promueve receptores a testosterona en cerebro y por otro inhibe el desarrollo de órganos femeninos ya que ciertas células del macho que están capacitadas para producir órganos femeninos al interactuar con la testosterona no pueden dividirse ni diferenciarse. Esto constituye un buen ejemplo de cómo la hormona (la cual es un producto del propio organismo) se comporta como una señal interna que causa que algunos genes se activen en ciertos tejidos embrionarios, cambiando la bioquímica y en última instancia el desarrollo de dichos tejidos. Estos efectos organizativos de la testosterona se dan en etapas anteriores al nacimiento y hace que las crías, al momento de nacer, sean machos o hembras tanto cerebral como orgánicamente (Gónadas). Luego de esta etapa organizativa los niveles de testosterona en sangre bajan, pero la hormona volverá a tener efectos importantes, ya no a nivel organizativo sino funcional, cuando el animal sea adulto.
En la rata hembra (sin el cromosoma "Y") no existe "Factor Determinante de Testículos" y por ende las células productoras de testosterona no se desarrollan y las células promotoras del ovario se dividen y diferencian dando origen a un ovario maduro, el cual producirá los estrógenos responsables del comportamiento femenino y sus ciclos de ovulación.
La testosterona posee dos efectos: Organizativo (Estructural): masculinización del cerebro (promueve receptores) cuando joven y de activación (Funcional): las hormonas hacen que las conductas sexuales se den ante estímulos adecuados.
Los efectos de las hormonas están limitados por factores temporales , por lo que una rata macho con cerebro de macho no responde a inyecciones de estrógenos (hormona femenina) aunque se le extraigan los testículos. El cerebro del macho sólo responde a la testosterona. Una rata hembra con un cerebro hembra, no se comporta como macho, aunque se le inyecte testosterona y se le saquen los ovarios. El cerebro hembra sólo responde a estrógenos.
Modelo epigenético esquemático
Un análisis de las interacciones recíprocas entre ambiente, genes e individuo tomando en cuenta los cambios en función del tiempo puede presentarse de la siguiente manera:
Si consideramos al cigoto como el estado original de un individuo (Fl), su fenotipo en el estado inmediato (F2) estará condicionado por los genes activos (Gl) que guíen el crecimiento y diferenciación durante un determinado período de tiempo y por el ambiente en el cual el desarrollo ocurra (Al). Así, cigoto + genes + ambiente darán origen al siguiente estado "F2": F1 + G1 + A1=F2.
El fenotipo en el estado siguiente "F3" estará determinado por la forma en que el fenotipo 2 (F2) cambie con un genoma "G2" (funcional en ese estado) y bajo un ambiente A2: F2 + G2 + A2 = F3.
Cada evento a lo largo del desarrollo establece un nuevo estado para el siguiente evento, pero NO rige sobre este.
En cuanto a Comportamiento Lorenz contribuyó a confundir el verdadero resultado del desarrollo comportamental, asegurando que ciertos comportamientos "Innatos" (Instinto) eran determinados directamente por los genes. Esta posición llevaba a una visión dicotómica donde había comportamientos innatos resultado de un programa fijo de desarrollo (denominado "blueprint") determinado por los genes y comportamientos aprendidos, los cuales eran adquiridos como resultado de la experiencia.
Esta visión fue duramente criticada por Lehrman (1970), Bateson (1981) y Hinde (1982) entre otros, tratando en todos los casos de resaltar las características epigenéticas de la ontogenia. En particular las críticas de Lehrman a la teoría de Lorenz permitieron reorientar el estudio etológico a un mayor énfasis en el desarrollo y dejando de lado la idea de instintivo por la de especie-específico.
De lo expuesto hasta aquí se puede concluir que para determinar la causa próxima (mecanismo) de que el comportamiento cambie durante el desarrollo debemos reconocer dos hechos concretos:
1. Las conductas NO pueden clasificarse como genética o ambientalmente determinadas .
2. En la búsqueda de los agentes de cambio durante el desarrollo debemos concentrarnos sobre el grado de influencia que los factores ambientales y la experiencia poseen sobre la ontogenia, pero no con una visión dicotómica sino planteando un continuo de variación en el grado de estabilidad del proceso de desarrollo frente a los factores externos .
Los agentes de cambio ontogénico
Siguiendo los dos conceptos anteriores, podemos reconocer diferentes agentes de cambio en el desarrollo conductual de un individuo.
1) La información genética:
El genoma contribuye activamente a que el desarrollo ontogenético siga ciertos patrones pre-organizados, característicos de grupos filogenéticamente relacionados (grupos emparentados, especies, etc.).
Dentro del genoma ciertos genes reguladores son los principales responsables en contribuir a la organización de los procesos de desarrollo embriológico. El genoma actúa diferencialmente dependiendo de patrones propios de expresión. A pesar de que el genotipo del individuo puede considerarse invariable a lo largo de su vida, para cada estadio del desarrollo, solo parte del genoma será funcional y es por esto que los modelos consideran distintos genotipos para cada etapa (G1, G2, etc.).
La actividad de los genes está controlada, entre otros factores, por la interacción de los genes entre si reconocida en fenómenos como:
ü Dominancia y Codominancia: interacción entre los alelos de un mismo gen, uno en cada homólogo.
ü Epistasia: un gen interfiere o modifica el efecto de otro gen.
ü Poligenia: un carácter está condicionado por más de un gen.
ü Pleiotropia: un solo gen afecta más de una característica.
Asimismo, hay un control de los estímulos externos sobre la actividad del genoma (fotoperiodos, hormonas, temperatura, interacción social, etc.).
Los genes producen proteínas reguladoras, las cuales no son efectivas de por sí, sino que deben encontrarse con un organismo sensible al mensaje que esta transporta. Dicha capacidad de dar respuesta funcional a esta proteína reguladora puede depender, entre otros factores, de la existencia de proteínas de la membrana celular denominadas receptores, capaces de interpretar dicho mensaje y dar una respuesta biológica, la cual consiste en determinar parámetros como: difusibilidad, adhesión celular; movilidad, tasas de proliferación celular, etc.
La organización interna del individuo en desarrollo condicionará la existencia del receptor, la cual puede depender de que el gen que lo produce sea funcional en esos momentos del desarrollo y que todos los elementos requeridos para la síntesis proteica estén presentes (aspectos ambientales y genéticos). Además, todo este intrincado mecanismo es viable, solo, bajo ciertas condiciones externas (pH, temperatura, radiación, etc.) a las cuales se suma el entorno celular como factor determinante del proceso de proliferación y diferenciación.
La complejidad del fenómeno de desarrollo en particular en alteraciones de la conducta humana y en relación al genoma, puede representarse con el ejemplo de la impronta genómica. Este fenómeno ocurre cuando la expresión de un gene depende de cuál de los dos padres le paso el gen al descendiente.
Según Alberch "un proceso de desarrollo embrionario, desde un punto de vista abstracto, es un proceso dinámico, que puede definirse como la emergencia progresiva de forma y heterogeneidad en base a la reiteración de una serie de reglas de interacción local, dentro de un contexto impuesto por unas condiciones iniciales".
El papel de los Genes en el desarrollo está relacionado con la determinación de las Leyes de Interacción y las Condiciones Iniciales del Sistema. La estructura final está condicionada por las interacciones dinámicas entre las leyes y los estados iniciales, pero una misma serie de reglas puede generar resultados diversos. Asimismo, se puede llegar a resultados finales semejantes partiendo de reglas diferentes. A pesar del dinamismo del sistema es importante destacar que la naturaleza misma de las reglas de interacción limita el número de resultados posibles.
2) El aprendizaje:
El aprendizaje consiste en un cambio más o menos permanente en el comportamiento que resulta de la experiencia con eventos ambientales. El aprendizaje no puede medirse en forma directa ya que representa un cambio en el potencial de respuesta. Lo medible es el desempeño del individuo y luego relacionarlo con un proceso de aprendizaje por parte de este. Asimismo, el aprendizaje es un proceso específico de manera tal que los cambios pueden darse para una conducta particular en una situación dada. Por lo tanto, el no observar cambios en la conducta no implica la falta de aprendizaje, tal vez pueda deberse a que la situación no es la indicada.
El aprendizaje se define por exclusión ya que los cambios en el comportamiento pueden deberse a otros factores como son la Maduración, la Fatiga, la Adaptación sensorial y la Motivación, por ende, debemos demostrar que ninguno de estos procesos se esté dando para considerar que le cambio es debido al aprendizaje.
No todos los cambios comportamentales que se ajustan a las características de un proceso de aprendizaje pueden ser considerados del mismo tipo. Una primera división permite clasificarlos en aprendizaje asociativo y no asociativo. A su vez el aprendizaje asociativo posee dos formas distinguibles elCondicionamiento Clásico y el Condicionamiento Operante. La diferencia principal entre ambos consiste en que en el Clásico (o Pavloviano) un evento importante es precedido por un estímulo (inicialmente neutro) el cual como resultado de la experiencia empieza a evocar la misma respuesta que el estímulo importante. En el caso del Condicionamiento Operante (o Instrumental) un evento importante sigue a una respuesta, y como resultado de ello hay un cambio en la probabilidad de dar dicha respuesta.
El aprendizaje no asociativo es una forma de aprendizaje simples, con cambios conductuales de corto tiempo, como la Habituación y Sensibilización. La Habituación consiste en la disminución en una respuesta debido a la exposición reiterada a un estímulo el cual no es seguido de ningún tipo de refuerzo
3) La Maduración:
Los procesos de maduración de los sistemas claves como el nervioso, hormonal, etc. condicionan el normal desempeño de la gran mayoría de las conductas para las cuales estos son mediadores o efectores.
4) La experiencia y dos Factores Claves del desarrollo:
Períodos Críticos y Estímulos Claves
Los efectos que un estímulo determinado posee sobre un mismo organismo en dos momentos de la vida pueden ser diferente. Este hecho está relacionado con las diferencias en sensibilidad, o cuan influenciable puede ser un individuo, en distintos períodos del desarrollo. Es aceptado que los estadios tempranos son más susceptibles a la experiencia y es posible identificar "períodos sensibles o críticos" en los cuales tanto una estimulación (interna o externa) como la propia práctica o actividad del individuo poseen efectos que cambian el estado de desarrollo de una manera pronunciada y muchas veces irreversible.
El modelo de "apego temprano" desarrollado principalmente en primates no humanos y humanos, constituye un buen ejemplo de cómo la interacción social en estadios tempranos del desarrollo posee efectos críticos sobre el desempeño comportamental general en las etapas inmediatas y posteriores del individuo, afectando diferentes aspectos como: social, cognitivo, emocional, etc. El aislamiento desde el nacimiento y durante seis meses posee efectos más marcados que el aislamiento durante períodos mayores a seis meses en etapas adultas de los individuos.
Kraemer propuso un modelo psicobiológico que relaciona los efectos del aislamiento social temprano en primates no humanos con alteraciones permanentes en ciertos sistemas de neurotransmisores relacionados con la regulación de respuestas ante la novedad (estrés). El sistema propuesto es el encargado de la síntesis cerebral de aminas biogénicas (neurotransmisores) entre las que poseen funciones claves: la noradrenalina, la dopamina y la serotonina; para las cuales se ha encontrado una desregulación importante como efecto del aislamiento social.
Concluyó que los efectos iniciales del aislamiento social se manifiestan en una reducción de la actividad del sistema de las aminas biógenas y una depresión comportamental. Esto hace que los sujetos no posean un sistema maduro que le permita dar respuestas "Normales" (más ajustada a nuevas situaciones que los ponen a prueba) ante situaciones novedosas. Un aislamiento social muy prolongado lleva a una "hipersensibilidad" tanto comportamental como neurobiológica cuando el sujeto es expuesto a condiciones novedosas.
Integración de agentes de cambio y el proceso ontogenético
La producción científica actual solo nos habilita a pensar que puede haber una correlación entre la presencia de una cadena particular de ADN (gen) y un rasgo. Asimismo, la mayor parte de los casos un gen no está relacionado causalmente con el rasgo y casi siempre el gen en cuestión no es una condición suficiente ni necesaria para el desarrollo del carácter.
Una forma interesante de entender que no hay una relación lineal causal entre genes y fenotipo, resulta de relacionar una idea intuitiva de “complejidad estructural y funcional de un organismo” con el número de genes encontrados al momento.
Existe un continuo de variación en el grado de estabilidad del proceso de desarrollo frente a los factores externos, cuando el proceso es estable a pesar de variaciones ambientales (no quiere decir que no cambia, sino que cambia siempre de la misma forma), el agente de cambio es la Maduración. En el otro extremo si modificaciones ambientales generan diversidad en los pasos del proceso entonces los agentes de cambios serán la experiencia y el aprendizaje. Existe una graduación intermedia de rasgos, la mayoría de los conocidos, donde se mezclan estos agentes.
Análisis de los Agentes de Cambio en el Desarrollo Individual:
1. No se pueden clasificar los comportamientos como innatos o aprendidos.
2. Debemos evaluar el grado de influencia que el ambiente y la experiencia
poseen sobre el curso del desarrollo individual.
Agente de Cambio
Estabilidad Alta
Maduración
Continuo
Estabilidad Baja Plasticidad
Aprendizaje y Experiencia
Períodos Críticos Estímulos Claves
Hay un continuo de variación en el grado de estabilidad del desarrollo frente a factores ambientales.
El desarrollo de un rasgo depende de una intrincada red entre los genes, sus productos y el ambiente.
Waddington propuso el paisaje epigenético para entender el proceso de desarrollo.
Por ejemplo, la presencia de estimulación con lenguaje de semejantes durante los primeros años de vida hace que el organismo en desarrollo se encamine al desarrollo normal. Luego dependiendo de la lengua con la cual se los estimule desarrollarán el idioma. Si fuera un niño sin estimulación suficiente para el lenguaje el sendero de desarrollo llevará a un protolenguaje. Una estimulación intensiva sólo dará algunas mejoras.
Los valles más profundos están asociados a rasgos que se denominan muy canalizados, los cuales muestran un patrón de cambios muy estable, poco bajo muy diversas condiciones ambientales y solo efectos muy pronunciados pueden provocar variaciones en el curso del desarrollo. Otros sectores del paisaje son más aplanados de manera que con el avance del desarrollo en el tiempo los patrones de cambio se modificaran producto de distintos factores que pueden alterar el rumbo y derivar en distintos productos, esos rasgos se dicen menos canalizados.
Así, poseer lenguaje es un rasgo muy canalizado, y solo condiciones ambientales extremas o alteraciones genéticas pueden alterar el normal desarrollo. El tipo de lengua dependerá del ambiente de crianza y el rasgo podrá cambiar sensiblemente, incluso en distintos momentos de desarrollo, aunque no tiene por qué ser equivalente la intensidad del cambio ambiental para lograrlo.
Los rasgos que dependen del agente de cambio maduración están muy canalizados, a medida que el aprendizaje y la experiencia cobran más importancia en el cambio del rasgo a lo largo del desarrollo, decimos que el rasgo esta menos canalizado. Como vemos la idea de genético o ambiental, desaparece conservando la idea de complejidad del proceso y respetando los agentes de cambio que intervienen.