La membrana plasmática está presente en todos los organismos vivos y define los límites celulares y constituye una barrera altamente selectiva que regula el intercambio de materiales con el entorno, permitiendo mantener el medio interno adaptado a las necesidades fisiológicas de cada organismo, así estableciendo una estrecha y controlada comunicación entre el interior y el exterior.
Las células eucariotas cuentan con un complejo sistema de membranas internas que delimitan compartimentos con procesos químicos y rutas metabólicas propias, un ejemplo de esto es el sistema de endomembranas o las membranas de organelas, como las mitocondrias, cloroplastos y peroxisomas.
Estas membranas son barreras semipermeables y dinámicas que tienen las siguientes funciones:
● regulan el tamaño y los límites de las células.
● Presentan una permeabilidad selectiva para regular el intercambio de sustancias entre el citoplasma y el medio extracelular y, además, entre los distintos compartimentos celulares de acuerdo con el tamaño y a las propiedades fisicoquímicas de las mismas.
● Delimitan medios que presentan composiciones y procesos químico-metabólicos distintos. Además de diferenciar el medio intra del extracelular, diferencian compartimentos celulares como el citosol del sistema de endomembranas o la matriz mitocondrial del espacio intermembrana.
● Permite el anclaje de las células con el medio extracelular.
● Facilita la interacción de las células con la matriz extracelular y las células vecinas. Esto gracias a receptores que desencadenan respuestas intracelulares.
Las células de todos los organismos vivos están delimitadas por membranas biológicas, estas responden a el modelo del mosaico fluido, ya que están formadas por una bicapa de fosfolípidos en la que se integran diversas proteínas como si fueran mosaicos. En medios acuosos, los fosfolípidos se organizan espontáneamente formando bicapas lipídicas compuestas por dos monocapas enfrentadas. En las membranas animales también está presente el colesterol.
Necesitaríamos una aclaración de la afirmación histórica de que la mb plasmática es la única que tiene oligosacáridos. La proporción de cada uno de estos componentes varía de acuerdo con las funciones de cada tipo de membrana. Todas las membranas biológicas, desde la membrana celular hasta la que rodea las estructuras membranosas intracelulares (mitocondrias, cloroplastos, peroxisomas y componentes del sistema de endomembranas, entre otros), están conformadas por: lípidos, proteínas e hidratos de carbono.
Lípidos: los principales que están presentes en las membranas son los fosfolípidos, los glucolípidos y el colesterol. Los fosfolípidos son los constituyentes que están más presentes en las membranas junto a las proteínas, estos lípidos son moléculas anfipáticas. Las colas carbonadas de los fosfolípidos pueden presentar dobles enlaces entre los átomos de carbono. En ese caso diremos que no están saturadas. En cambio, si no presentan dobles enlaces diremos que están saturadas. Sus cabezas van hacia el medio acuoso y las colas no polares hacia el interior hidrofóbico de la bicapa. En un medio acuoso las bicapas tienden a replegarse sobre sí mismas y a formar espontáneamente compartimientos cerrados como vesículas o incluso células. El interior hidrofóbico de la bicapa genera una barrera que impide el pasaje de la gran parte de las sustancias hidrofílicas, pero no el de las hidrofóbicas. En las membranas de células animales encontramos otro lípido exclusivo de estas células, el colesterol, que está ausente en el resto de los organismos vivos,se ubica entre los fosfolípidos. Tanto la presencia de colesterol como la saturación de las colas de los fosfolípidos permiten regular la fluidez de las membranas.
Proteínas: son responsables de muchas de las funciones específicas de las membranas, cada una tiene una composición proteica distintiva. Las proteínas de membrana pueden asociarse a la bicapa lipídica de distintas maneras:
● Proteínas integrales o intrínsecas: pueden atravesar parcial o totalmente la bicapa. En este caso se las denomina transmembrana. Se diferencian en monopaso y multipaso.
● Proteínas periféricas: se adosan a las membranas desde el exterior o el interior de las células.
Hidratos de carbono: Son mucho más abundantes en la membrana plasmática, preferentemente localizados en la monocapa externa. Se ubican en la cara extracelular de la membrana plasmática formando una estructura llamada glicocálix y se unen tanto a lípidos como a proteínas. Entre sus funciones más importantes podemos citar su participación en procesos de reconocimiento celular de sustancias ajenas al organismo y de moléculas propias del cuerpo. Los oligosacáridos que forman parte de receptores brindan al organismo una identidad propia que nos diferencia unos de otros.
Propiedades de las membranas
De acuerdo con el modelo del mosaico fluido, las membranas son fluidos bidimensionales, dinámicos y viscosos, cuyas moléculas están siempre en movimiento. A continuación, vamos a desarrollar las características de la fluidez de las membranas:
● Los fosfolípidos pueden presentar distintos tipos de movimientos. Algunos movimientos suceden de manera espontánea, como la difusión lateral y la rotación. Otros como el flip flop no ocurren libremente debido a la dificultad de las cabezas polares de traspasar el medio hidrofóbico interno de la membrana. Las proteínas integrales también pueden difundir y rotar sobre su propio eje, pero no girar a través del plano de la membrana. Suelen desplazarse acompañadas de los lípidos que las rodean.
● Existen factores que pueden afectar a la fluidez de las membranas. La variación en la temperatura es el primer factor, al incrementar la temperatura aumenta el movimiento propio de moléculas, como los fosfolípidos, y por ende se incrementa la fluidez. En cambio, una disminución en la temperatura hace más rígidas las membranas. La presencia de ácidos grasos insaturados es el segundo factor, cuanta más presencia de colas insaturadas hay en los fosfolípidos, mayor será la fluidez de la membrana, ya que la insaturación de colas genera un acodamiento que restringe la compactación entre ellos. En cambio, si hay una mayor saturación se facilita la compactación de colas y disminuye la fluidez. Por último, el tercer factor es la presencia de colesterol, regula el grado de fluidez, a los 37º C restringe el movimiento excesivo de los fosfolípidos, así disminuyendo la fluidez y estabilizando las membranas. A menos de 37º C se evita que los fosfolípidos se compacten demasiado y aumenta su fluidez.
● Las membranas son asimétricas. Sus composiciones son distintas dado que los procesos que se desarrollan sobre cada una de las caras también lo son. En el caso de la membrana plasmática, los glicolípidos y las glicoproteínas, implicados en el reconocimiento celular, se ubican casi exclusivamente en la cara expuesta hacia el medio extracelular.
● Las membranas biológicas presentan permeabilidad selectiva. La membrana plasmática actúa como una barrera selectivamente permeable que regula el tránsito de moléculas y mantiene así el medio interno celular dentro de parámetros relativamente constantes. Permite la difusión de moléculas no polares siempre y cuando su tamaño no sea muy grande. Sin embargo es impermeable a la mayor parte de moléculas polares. El resto de las moléculas deberá ser transportado de un lado a otro de la membrana por medio de proteínas integrales que actúan como transportadores.
Transporte a través de las membranas
Podemos definir a la difusión como el movimiento pasivo y sin gasto de energía de moléculas desde una zona de mayor concentración hacia una que tiene menos concentración. Y al gradiente como la diferencia de concentraciones entre ambas zonas. Cuando una sustancia se desplaza sin gasto de energía desde zonas de mayor concentración hacia zonas que tienen menos concentración decimos que difunde a favor del gradiente de concentración, en cambio cuando el movimiento es en contra del gradiente se requerirá de un aporte energético.
Cuando las moléculas se transportan sin gasto de energía a favor de su gradiente el desplazamiento se considera transporte pasivo. Pero si este movimiento se lleva a cabo en contra del gradiente y con gasto de energía se lo llama transporte activo. El transporte en masa, en cambio, es independiente del gradiente de las sustancias a transportar
Transporte pasivo
Difusión simple. Es el movimiento de sustancias entre compartimientos separados por una membrana permeable a las mismas y sin participación de proteínas transportadoras. Como el proceso es a favor del gradiente no requiere de un aporte de energía.
● Se le dice osmosis al caso del movimiento del oxígeno, dado que las membranas son relativamente permeables al agua, pero impermeables a muchos solutos, el h2o difundirá desde el compartimiento con menor concentración de solutos o hipotónica al de mayor concentración, o hipertónica para equilibrar ambas concentraciones. Si una célula se coloca en una solución cuya concentración es similar al interior celular es una solución isotónica. Si la célula se sumerge en una solución hipertónica más concentrada que el interior celular, el agua tenderá a salir y la célula se va a deshidratar.
Finalmente, si la célula se coloca en una solución hipotónica, menos concentrada, el agua ingresa a la célula, lo que puede llevar a su lisis.
Difusión facilitada. Las bicapas lipídicas son altamente impermeables a todos los iones y a la mayoría de las moléculas polares sin carga. En consecuencia, estas sustancias necesitarán de proteínas específicas que permitan translocar las de un lado a otro de la membrana. La difusión facilitada ocurre siempre a favor del gradiente y por ello no requiere de un aporte de energía. Sin embargo, el movimiento puede depender solo de un gradiente químico (diferencias de concentración) si no hay cargas involucradas, o de un gradiente electroquímico (diferencias de concentración y de carga) si se trata de moléculas con carga. Podemos diferenciar dos tipos de proteínas que participan en la difusión facilitada:
● Canales iónicos. Son proteínas altamente selectivas que facilitan el paso de iones a una velocidad elevada. Están presentes en todas las membranas, tanto en la plasmática como en las de los organoides y, en general, transportan iones de tamaño pequeño. Algunos canales requieren para su apertura de estímulos externos, por ejemplo, la presencia de una molécula inductora (canales dependientes de ligando) o una modificación de la carga eléctrica a ambos lados de la membrana (canales dependientes de voltaje). El transporte de un ion es impulsado por su gradiente electroquímico. Es decir que la fuerza que impulsa un ion a través de la membrana se debe tanto a su diferencia de concentración como a la diferencia de cargas a ambos lados de la membrana.
● Proteínas “carrier” o transportadores. Son proteínas específicas que transportan una gran variedad de moléculas polares a favor de su gradiente y no requieren un gasto de ATP. A diferencia de las proteínas de canal, los carriers sufren un cambio conformacional durante el pasaje de moléculas. Las etapas de este transporte son las siguientes:
1. La proteína se une específicamente al soluto.
2. Sucede un cambio conformacional reversible que permite transportar el soluto de un lado al otro de la membrana.
3. El soluto se libera y el carrier vuelve a su forma original. Cabe aclarar que este tipo de transporte es mucho más lento que el que se realiza a través de canales.
Transporte activo
Las células necesitan mantener su homeostasis incorporando algunas sustancias y eliminando otras, independientemente de su concentración en el medio externo. Para ello cuentan con mecanismos que permiten transportar moléculas en contra de su gradiente electroquímico, con gasto energético. Una gran parte de la energía de la célula se destina para estos procesos de los que participan proteínas llamadas bombas. Ya que este transporte se desarrolla en contra del gradiente, las bombas requieren de la energía que aporta el ATP, la “moneda energética” celular. Se puede subdividir en:
● Transporte activo primario (acoplado directamente a la ruptura del ATP). Uno o más solutos son bombeados en contra de su gradiente electroquímico gracias a la energía aportada por el ATP.
● Transporte activo secundario (acoplado a gradientes generados por una bomba) Una sustancia B deberá ser transportada en contra de su gradiente. Pero no se usará directamente una bomba para el transporte, sino que se aprovechará el gradiente de otra sustancia A (puede tratarse de Na + o de H + ) bombeada previamente con gasto de ATP. En conclusión, el gradiente iónico de A será utilizado para transportar en contra de su gradiente otro soluto (B), que de otra manera no podría lograrlo.
Las proteínas transportadoras se pueden clasificar de acuerdo a la cantidad de solutos que transportan y su dirección:
-Uniporte: transfieren UN solo tipo de soluto de un lado al otro de la membrana.
-Simporte: transfieren DOS tipos de solutos, ambos en la misma dirección.
-Antiporte: transfieren DOS tipos distintos de solutos en sentidos contrarios.
● Transporte activo primario: Bomba Na+K+
Esta concentración desigual a ambos lados de las membranas se mantiene gracias a la acción de la Bomba Na + -K + que está presente en todas las membranas plasmáticas de las células animales y tiene función de transporte y de ATPasa. La bomba transporta 3 átomos de Na+ al citosol. Ambos iones son movilizados en contra de sus gradientes electroquímicos, lo que permite generar las diferencias de concentración y carga entre ambos espacios. Es importante recordar que, ambos iones se transportan en contra de su gradiente y por eso se necesita ATP.
● Transporte activo secundario.
Este tipo de transporte se denomina transporte activo secundario ya que hay un compuesto que se mueve en contra de su gradiente y requiere, de manera indirecta, el gasto del ATP de la bomba para generar un gradiente de Na+, cuyo potencial electroquímico permita el ingreso conjunto con la glucosa
Transporte en masa
Se requiere de la participación de una porción más extensa de membrana así formando vesículas. Si se incorporan sustancias que provienen del exterior hablamos de una endocitosis. Si las sustancias abandonan las células se trata de una exocitosis. Este transporte es independiente del gradiente electroquímico y requiere siempre de un gasto del ATP y la participación del citoesqueleto.
● Endocitosis. Es un proceso en el cual un sector de la membrana rodea progresivamente al material que será internalizado a la célula hasta que queda englobado en una vesícula endocítica. Se pueden distinguir 3 tipos de procesos de acuerdo con el tipo de sustancia que va a entrar a la célula:
1. Fagocitosis: Es un proceso en el cual ciertas células rodean con su membrana a partículas sólidas, restos celulares o microorganismos y las incorporan al interior celular. Esto se desarrolla gracias a la emisión de pseudópodos que engloban la partícula hasta finalmente incorporar a la célula en forma de vesícula llamada fagosoma. Luego, se fusiona con otra vesícula para formar los lisosomas.
2. Endocitosis mediada por receptor: es un proceso muy selectivo donde los receptores específicos de la membrana plasmática reconocen a moléculas que luego serán endocitadas en forma de una vesícula llamada endosoma.
3. Pinocitosis. Es la incorporación inespecífica de líquidos y moléculas disueltas en él a través de vesículas de tamaño reducido.
● Exocitosis. Es el proceso donde las vesículas provenientes del aparato de golgi se fusionan con la membrana plasmática y el material que está contenido en las mismas es liberado al medio extracelular. La membrana de la vesícula se integra a la membrana plasmática. La secreción de sustancias puede ser continua, donde las moléculas son liberadas sin estímulos externos, pero también regulado, donde señales externas a la célula inducen a liberar contenido de las vesículas de secreción.
Las membranas delimitan distintos espacios intracelulares
Las células eucariontes poseen estructuras membranosas internas que permiten generar espacios o compartimentos intracelulares diferenciados, cada uno con una función característica. En estos compartimientos membranosos se lleva a cabo una gran variedad de procesos metabólicos, muchos de ellos relacionados con transformaciones energéticas, así como con la síntesis y degradación de biomoléculas.
Podemos diferenciar a grandes rasgos tres tipos de estructuras membranosas:
● La membrana plasmática, aquella que delimita el interior citoplasmático del exterior celular, sea de otras células en el caso de organismos pluricelulares o del ambiente donde vive ese organismo en el caso de organismos unicelulares.
● Membranas de organelas membranosas, aquellas que delimitan estructuras como mitocondrias, cloroplastos y peroxisomas. Cabe aclarar que los procariotas no cuentan con organelas membranosas internas. Tanto las membranas de las mitocondrias como las de los cloroplastos presentan un complejo conjunto de proteínas integrales involucradas en procesos de transformación energética. En el caso de los cloroplastos, que llevan a cabo la fotosíntesis, los complejos proteicos y la clorofila ubicados sobre sus membranas permiten la transformación de energía lumínica en energía química (síntesis de ATP). Por otro lado, gran parte de la respiración celular se desarrolla sobre las membranas mitocondriales, donde la energía contenida en las biomoléculas es utilizada para sintetizar el ATP necesario para todos los procesos celulares que requieran de energía.
● Las membranas que conforman el sistema de endomembranas o sistema vacuolar citoplasmático (SVC). Son estructuras membranosas interrelacionadas presentes en organismos eucariontes, con distintas funciones, una de ellas consiste en facilitar el ingreso de estos glúcidos a las células para así degradarlos y obtener energía o para sintetizar polisacáridos como el glucógeno. Se trata de un “sistema” dado que sus componentes están relacionados y trabajan de modo interdependiente.
En el caso de las células eucariontes, este sistema de membranas internas permite separar procesos metabólicos en distintos compartimentos, generando las condiciones que cada proceso requiere y permitiendo a la vez una fluida interconexión con el resto de las estructuras y procesos celulares.
El Sistema Vacuolar Citoplasmático se conforma por las siguientes componentes, aunque cada aclarar que este sistema no pertenece a organelas como las mitocondrias, cloroplastos ni los peroxisomas a pesar de estar rodeados y conformados por membranas. La membrana plasmática tampoco pertenece a este sistema, si bien existe un transporte continuo y fluido entre ambas estructuras.
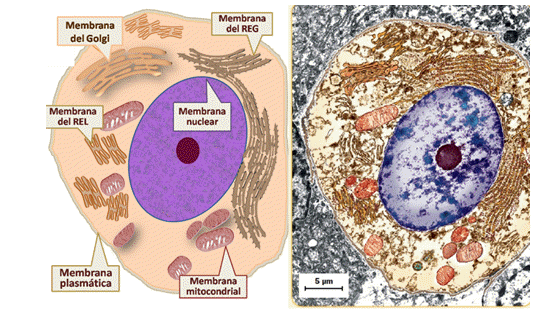
1. La envoltura o membrana nuclear o carioteca: rodea y contiene al material genético de la célula y presenta poros que permiten el transporte de sustancias desde el citosol hacia el interior del núcleo y viceversa. Sus membranas presentan continuidad con la del REG y su membrana externa presenta ribosomas adheridos sobre su cara citoplasmática.
2. Los retículos endoplasmáticos: se divide en REL (retículo endoplasmático liso) y RER (retículo endoplasmático rugoso). El primero (REL) está constituido por túbulos membranosos delgados que carecen de ribosomas. Desempeña las siguientes funciones:
● Síntesis de lípidos, entre ellos esteroides como el colesterol, fosfolípidos y triglicéridos. Es el responsable de la síntesis del componente lipídico de las membranas biológicas.
● Detoxificación de sustancias como el alcohol, principalmente en células del hígado.
● Degradación del glucógeno.
● Reservorio de calcio. Presenta bombas de Ca 2+ que transportan este ión desde el citosol al lumen del REL. Esto es particularmente importante en las células musculares. El Ca 2 + es un importante mensajero intracelular que es liberado al citosol en respuesta a un estímulo específico.
El RER es un sistema de sacos membranosos interconectados con el REL y con la membrana nuclear. Presenta ribosomas adheridos en su cara citosólica, su presencia se relaciona con su función principal: la síntesis y glicosilación de las siguientes proteínas:
● Las proteínas de membrana.
● Las proteínas de exportación.
● las enzimas que pertenecen al SVC.
● Las enzimas hidrolíticas o lisosomales.
3. El complejo o aparato de Golgi: Está constituido por distintas cisternas (o sacos membranosos) apiladas, que se subdividen en tres zonas: la cis (más cercana al REG o al REL), la medial y la trans (más cercana a la membrana plasmática). Algunas de sus funciones son: el procesamiento de biomoléculas que se sintetizaron en el REL y el procesamiento de las proteínas que se sintetizan en el REG.
4. Lisosomas: Se trata de orgánulos con una alta concentración de enzimas hidrolíticas que tienen a su cargo la degradación de las macromoléculas derivadas de los procesos endocíticos así como de componentes propios de la célula.
5. Vesículas de transporte intracelular: La función de estas vesículas es el transporte de sustancias entre distintas estructuras membranosas.
6. Endosomas: Se trata de vesículas formadas a partir de los distintos procesos endocíticos.
Transporte de sustancias entre compartimentos del SVC
El transporte de sustancias entre compartimentos se lleva a cabo a través de vesículas, que son pequeñas bolsas membranosas. Se generan por brotación a partir de la membrana de un compartimiento generador, se transportan por el citosol y luego se funden con la membrana aceptora. Luego de ser procesadas, abandonan el compartimiento transportadas nuevamente por vesículas y se dirigen, en caso de tratarse de proteínas de secreción, a la membrana plasmática donde son liberadas al espacio extracelular gracias a la fusión de la vesícula con la membrana.
Síntesis y direccionamiento de lípidos:
Los lípidos se sintetizan en la membrana del REL, continúan en la membrana del Golgi y finalmente son transportados en vesículas hacia la membrana plasmática de la que formarán parte.
Tráfico y direccionamiento intracelular de proteínas:
El REG es el lugar de síntesis de distintas proteínas celulares y extracelulares. Muchas serán posteriormente glicosiladas por agregado de oligosacáridos que luego se modificarán y terminarán de procesar en el Golgi. Desde allí son transportadas por medio de vesículas hasta su ubicación final, que pueden ser:
● Los lisosomas, en caso de tratarse de enzimas hidrolíticas.
● La membrana plasmática, si se tratara de proteínas de membrana (las bombas, los carriers y/o los receptores).
● La matriz extracelular, en el caso de proteínas de secreción (por ejemplo, hormonas como la insulina que serán transportadas luego por vía sanguínea o componentes que forman la matriz extracelular como el colágeno).
Digestión celular y lisosomas:
Son organelas especializadas en la digestión intracelular controlada, tanto de materiales extracelulares incorporados por endocitosis, como de organelas propias de las células y de otras biomoléculas. Las mismas contienen en su interior enzimas que hidrolizan las uniones covalentes de distintas biomoléculas. A estas enzimas, se las denomina hidrolasas ácidas ya que actúan a pH ácido. Por ello, el interior de los lisosomas presenta un pH de alrededor de 5. Las vesículas que brotan del complejo de Golgi contienen enzimas hidrolíticas, pero todavía carecen de las sustancias a degradar. Cuando se fusionan con vesículas conteniendo el material a degradar, se denominan lisosomas o vesículas digestivas y hay tres tipos de vías que conducen a su formación.
1. La endocitosis mediada por receptores. En la vesícula endocítica, el endosoma se fusiona con una vesícula con enzimas hidrolíticas proveniente del Golgi y se forma así el lisosoma.
2. La heterofagocitosis. que incorpora sustancias particulares como bacterias desde el exterior de la célula. Se forma así una vesícula endocítica denominada heterofagosoma y que finalmente también llevará a la formación de un lisosoma.
3. La autofagocitosis. La vesícula formada se denomina autofagosoma. En este proceso, organelas como las mitocondrias son degradadas por presentar algún daño o porque la disponibilidad de energía de la célula es insuficiente. En consecuencia, la célula degradará sus propias estructuras y biomoléculas contenidas en ellas para obtener energía.
Acá se ejemplifica el proceso fagocítico de una bacteria y su posterior degradación en un lisosoma por parte de un glóbulo blanco. Como producto de esta digestión celular, las estructuras bacterianas, la membrana lipídica, el ADN y los ribosomas son degradados totalmente. Las biomoléculas de la bacteria aprovechables son incorporadas por la célula. En cambio, las sustancias que no se pueden degradar son eliminadas por medio de vesículas secretoras hacia el exterior de la célula.
Las células y el medio (machete)
El citoplasma: Es el espacio comprendido entre la membrana plasmática y el núcleo (en eucariotas) o el nucleoide (en procariotas). En eucariotas está conformado por una región fluida, denominada citosol y por una serie de estructuras como los ribosomas, las organelas, el citoesqueleto y el sistema de endomembranas. El citoplasma no es estático, sino que está en constante movimiento. Lo conforman:
● Citosol. Es una sustancia acuosa semifluida que ocupa los espacios entre las organelas. Está compuesto por agua y sustancias disueltas. En él ocurren gran parte de las reacciones químicas de la célula.
● Ribosomas. Se los encuentra libres en el citosol o bien adheridos al retículo endoplasmático rugoso.
● Sistema de endomembranas. sistema de Golgi, retículos, lisosomas y vesículas.
● Organelas membranosas. Mitocôndrias, cloroplastos, peroxisomas.
● Citoesqueleto. Está conformado por una red de filamentos proteicos.
El citoesqueleto: Cada célula de un organismo cumple una función especializada. Algunas, como las células musculares, tienen la capacidad de contraerse. Otras, como los glóbulos blancos o los espermatozoides, pueden incluso desplazarse. Para esto se requiere de un conjunto dinámico de filamentos proteicos que faciliten estos procesos. El citoesqueleto cumple esta función y se conforma por distintos tipos de fibras: los microtúbulos, los microfilamentos y filamentos intermedios. Entre sus funciones básicas podemos nombrar:
-Dar forma y estabilidad a la célula
-Permitir el desplazamiento de ciertas células
-Transporte intracelular de organelas, vesículas, ribosomas, etc.
-Participar de la contracción muscular.
1.Los microtúbulos son estructuras en forma de tubo hueco que pueden alcanzar grandes longitudes. Están conformados por una proteína globular, la tubulina, que puede asociarse a otras tubulinas y formar así los microtúbulos. Pueden presentar formas transitorias o estables. Los microtúbulos transitorios tienen la capacidad de polimerizarse (armarse) o depolimerizarse (romperse o desarmarse) de acuerdo con los requerimientos de la célula. Esto es común en microtúbulos citoplasmáticos cuya función es actuar como un sistema de transporte intracelular, tanto en interfase como durante la división celular. En los microtúbulos estables, en cambio, 2 o 3 microtúbulos se asocian entre sí para formar estructuras con mayores dimensiones, presentes tanto en centríolos, cuerpos basales como en la estructura interna de cilios y flagelos. La función de los microtúbulos es:
● Transportar intracelularmente las vesículas, ribosomas, mitocondrias, etc.
● Transporte de cromosomas durante la división celular por medio de la formación del huso mitótico.
● Forma parte de la estructura de cilios, flagelos, cuerpos basales y centriolos.
-Centríolos y cuerpos basales: Los centríolos, corresponden a dos estructuras tubulares enfrentadas formadas por 9 tripletes de microtúbulos periféricos. La notación de 9+0 se refiere a los 9 tripletes periféricos, y el 0 se refiere a la ausencia de microtúbulos al interior del tubo. Los centríolos están inmersos de una matriz amorfa: el centrosoma. Los cuerpos basales, por otro lado, se conforman por una sola estructura tubular y se ubican bajo la membrana plasmática, en el nacimiento de cilios y flagelos, siendo los responsables de su formación.
-Flagelos y cilios: Los flagelos son largas prolongaciones de la membrana plasmática que permiten el desplazamiento de las células, como en el caso de los espermatozoides. Los cilios tienen una estructura similar, pero más corta. Ambos presentan 9 pares de microtúbulos organizados formando un círculo y un par adicional al centro del anillo. Esta estructura se conoce como disposición 9+2.
2.Los microfilamentos están constituidos por una proteína llamada actina G que a su vez puede asociarse con otras actinas y formar así largos filamentos. Se trata de estructuras dinámicas que se forman y rompen rápidamente de acuerdo con las necesidades de la célula. Entre sus funciones se encuentran:
● Dar fórmula a las células
● participar de la contracción muscular
● colaborar durante la división celular en la separación de las células hijas
● permitir el desplazamiento celular gracias a las prolongaciones celulares
● participar directamente en procesos de endocitosis y exocitosis
● formar parte de microvellosidades
3.Los filamentos intermedios brindan resistencia interna a las células. La proteína que los constituye, al ser de naturaleza fibrosa y no globular, permite esta fortaleza mecánica. Los filamentos intermedios, más precisamente los conformados por queratina, también participan de diferenciaciones de la membrana plasmática denominadas desmosomas y hemidesmosomas y que están presentes en algunos tipos celulares.
Comunicación entre células
Todo organismo vivo debe ser capaz de reconocer el ambiente que lo rodea, detectando cambios que se generen en el mismo y respondiendo a ellos. En los organismos pluricelulares, la información proveniente de células vecinas o del exterior a su vez debe ser transmitida a otras células del organismo y coordinada entre las mismas, por lo que son necesarios distintos sistemas de señalización o comunicación celular. Estos sistemas de comunicación son imprescindibles ya que controlan tanto el crecimiento, el funcionamiento como también la organización de las células en los tejidos y órganos. Es decir, esa comunicación entre células se requiere para el correcto funcionamiento de todo el organismo.
● Célula secretora o inductora: sintetiza y libera una molécula señal o ligando, muchas veces en respuesta a un estímulo externo o interno. Estas células sintetizan las moléculas en el sistema de endomembranas (retículo endoplásmico o RE, Golgi) y luego la señal es liberada hacia el medio extracelular.
● Ligando, señal o inductor: molécula sintetizada por la célula inductora, que genera un efecto o respuesta en la célula inducida, es decir en aquella que presenta los receptores específicos para ese ligando o señal. De este modo iniciará una respuesta celular concreta. Célula diana o inducida: es la célula a la que se une el ligando y es la que presenta los receptores específicos que reconocerán a la molécula señal. Esta célula puede encontrarse cercana o lejana a la célula inductora.
● Receptor: molécula a la que se une específicamente el ligando. Puede estar ubicado tanto en el citoplasma como en la membrana plasmática de la célula diana. La unión a este receptor por parte del ligando genera una respuesta específica celular.
Los ligandos o señales pueden recorrer, de acuerdo con el tipo de proceso que se trate, una distancia más o menos grande desde la célula inductora hasta la célula inducida. A continuación, se ejemplifican algunas de estas vías:
a) Vía parácrina: el ligando liberado por la célula secretora se une a receptores de las células que se encuentran en las inmediaciones. Se trata de un mecanismo de “diálogo” entre células vecinas. En este caso la distancia que recorre el ligando hasta la célula inducida es corta.
b) Vía endócrina: el ligando puede recorrer largas distancias desde su liberación hasta llegar a la célula diana. La vía de transporte suele ser la sanguínea. Los ligandos de esta vía se denominan hormonas. Ejemplos de hormonas serían la insulina, la testosterona, los estrógenos y las hormonas tiroideas, entre otras. Este mecanismo permite a las señales llegar a un gran número de células que se encuentran distantes de aquella que sintetizó la señal química.
c) Vía neuronal (sinapsis):. Este proceso lo llevan a cabo células especializadas llamadas neuronas que emiten largas prolongaciones citoplasmáticas. Los ligandos de esta vía, denominados neurotransmisores, se liberan a un espacio muy reducido que separa una neurona de la siguiente, en la cual se ubican los receptores.
d) Vía autócrina: el ligando generado y liberado por la célula secretora se une a los receptores de la misma célula. Habitualmente se trata de mecanismos de autorregulación que le permiten a la célula cuantificar la cantidad de ligando presente en el medio.
Las señales químicas pueden ser hidrofílicas o hidrofóbicas:
Los ligandos pueden ser tanto de naturaleza hidrofílicas (proteínas, glicoproteínas) como hidrofóbicas (lípidos, gases) y esto determina la ubicación de los receptores.
Señales hidrofóbicas: Atraviesan la membrana plasmática por difusión pasiva e ingresan al interior de la célula, donde se unen a su receptor específico.
Señales hidrofílicas: Estas señales no pueden atravesar la bicapa lipídica y por lo tanto su receptor se ubicará sobre la membrana plasmática.
En el caso de algunos ligandos hidrofóbicos de tipo hormonal como la testosterona, el ligando ingresa a la célula y allí se une al receptor y juntos ingresan al núcleo. Este receptor presenta una función específica: activar la expresión de ciertos genes, es decir, inducir la síntesis de proteínas.
En el caso de los ligandos hidrofílicos, la unión del ligando al receptor genera, como respuesta, una molécula señal intracelular. Este proceso, en donde un tipo de señal (extracelular) activa la síntesis de otra (intracelular), implica una “transducción de señal”. Esta señal intracelular, a su vez genera otras señales, encadenadas sucesivamente entre sí. Esto permite la amplificación de la señal, dado que en cada paso, una molécula induce la formación de múltiples nuevas moléculas. La respuesta celular debida a un ligando hidrofílico podría llevar a una modificación de la actividad tanto de proteínas intracelulares como de proteínas del citoesqueleto. Pero también pueden conducir a cambios en la expresión de los genes, estimulando o inhibiendo la síntesis de nuevas proteínas. En algunas vías de transducción relacionadas con ligandos hidrofílicos intervienen intermediarios químicos intracelulares denominados segundos mensajeros. Entre ellos podemos nombrar el AMPc (AMP cíclico) y el Ca 2+ (calcio).
Matriz extracelular
Es la sustancia que ocupa el espacio ubicado entre las células. No existe una matriz extracelular definida, sino que la misma varía de acuerdo a cada tipo de tejido. Presenta una función mecánica y estructural pero también se relaciona con la regulación de la forma y funciones celulares (como la proliferación, la migración de las células y desarrollo embrionario). Sus componentes son:
1. Proteoglucanos. Son la base fundamental de la matriz extracelular. Se trata de sustancias conformadas por proteínas e hidratos de carbono, muy ricas en cargas negativas. Por esta razón estas moléculas atraen agua y en consecuencia están muy hidratadas. Forman geles muy hidratados que funcionan del mismo modo que una esponja embebida en agua: si reciben presión, se deforman y expulsan el agua. Si dejan de recibir presión, recuperan la forma original y se rehidratan.
2. Proteínas fibrosas. Son proteínas que están inmersas en la matriz de proteoglucanos:
● El colágeno brinda a la matriz resistencia a la tracción. Es una molécula muy resistente formada por tres cadenas polipeptídicas unidas entre sí por puentes de hidrógeno. Su síntesis se lleva a cabo en el Retículo Endoplasmático Rugoso y se modifica en el Golgi, pero su maduración se da en la matriz extracelular.
● Elastina le da a la matriz propiedades elásticas, es decir que ante tensiones puede deformarse, pero cuando la tensión cesa, recupera su forma original.
3. Proteínas de adhesión. Son proteínas que forman parte de la matriz extracelular y posibilitan la unión de ésta con las células.